Биология для выпускников школ и поступающих в вузы - Мустафин А. Г. 2015 год
НАСЛЕДСТВЕННОСТЬ - ОБЩАЯ БИОЛОГИЯ
Генетика — изучает наследственность и изменчивость — универсальные свойства живого. Наследственность — способность организмов передавать свои признаки и особенности развития потомству. Благодаря наследственности все живые существа сохраняют в своих потомках характерные черты вида. В основе наследственности лежат особенности строения и функций нуклеиновых кислот клеток.
Геном, ген. Развитие представлений о гене
Единицей наследственной информации является ген — участок (локус) хромосомы, обеспечивающий возможность развития признака организма. Экспрессия генов в процессе развития организмов приводит к синтезу белков, дифференцировке клеток, формированию тканей и органов, становлению фенотипа — совокупности всех признаков организма.
Геном — это вся ДНК в гаплоидном наборе хромосом данного вида. В часности, размер генома человека составляет около 9 млрд пар нуклеотидов ДН К. По результатам выполнения проекта “Геном человека” только 1% ДНК кодирует синтез белков, 24% приходится на некодирующие интроны и 75% — на межгенные промежутки. Всего в геноме человека содержится 20 000—25000 генов. В процессе оплодотворения происходит объединение геномов мужского и женского организмов в единую систему — генотип. Генотип — совокупность генов, имеющих фенотипическое проявление в соматических клетках диплоидного организма. Гены, расположенные в одинаковых локусах гомологичных хромосом и отвечающие за развитие вариантов какого-либо признака, называют аллельными. Их принято обозначать буквами латинского алфавита. Аллельные гены могут быть доминантными (А, В) или рецессивными (а, b).
Организм, в локусах гомологичных хромосом которого находятся разные аллели (Аа; АаВb), называют гетерозиготой. Если же в соответствующих локусах гомологичных хромосом расположены одинаковые аллели (АА, аа; ААВВ, aabb), то такой организм называют гомозиготой по одному или нескольким признакам.
Доминантный аллель обусловливает развитие признака как у гомозигот, так и у гетерозигот. Рецессивные аллели вызывают развитие признака только в гомозиготном состоянии. Разные аллельные формы генов возникают в результате мутации — изменения структуры полинуклеотидной последовательности ДНК. Ген может мутировать много раз, образуя много аллелей. Если в генофонде популяции существует серия мутаций какого-либо гена, определяющая многообразие вариантов признака, то имеет место явление множественного аллелизма. Примером множественных аллелей может служить наследование групп крови человека по системе АВО, детерминируемых геном I (табл. 7.1). В популяциях людей существуют три аллеля гена; I°, Ia, Ib. В генотипах индивидуумов они комбинируются попарно, кодируя антигены поверхности эритроцитов.
Таблица 7.1
Группы крови человека в системе АВО
|
Аллели гена I |
Возможные генотипы |
Частота у населения (средние данные среди населения стран Европы), % |
|
I° |
I°I° |
46 |
|
IA |
IAIA; IAI° |
42 |
|
IB |
IBIB; IBI° |
9 |
|
IAIB |
3 |
Обычно исследователи имеют дело не непосредственно с генами, а с результатами их проявлений признаками или свойствами организма. При изучении закономерностей наследования признаков рассматривают два организма, являющихся партнерами при скрещивании. Обычно скрещивают организмы, отличающиеся контрастными альтернативными вариантами проявления какого-либо признака. Совокупность всех признаков организма на определённой стадии онтогенеза называют фенотипом. Фенотип формируется в процессе реализации наследственной информации генотипа под воздействием факторов окружающей среды.
Моногибридное скрещивание. Первый и второй законы Менделя
Гибридологическим метод, заключающийся в скрещивании и последующем учете расщеплений (соотношений фенотипических и генотипических разновидностей потомков), был разработан Г. Менделем (1865). К особенностям этого метода относят:
• прослеживание при скрещивании не всего многообразного комплекса признаков у родителей и потомков, а анализ наследования отдельных, выделяемых исследователем альтернативных признаков. Моногибридное скрещивание — прослеживают наследование одного признака, дигибридное — двух, полигибридное — нескольких признаков;
• количественный учет в ряду последовательных поколений гибридных потомков, различающихся по отдельным признакам:
• индивидуальный анализ потомства каждого организма;
• Г. Мендель использовал для экспериментов чистые линии, т.е. совокупность гомозиготных по одному или нескольким признакам организмов, при скрещивании которых между собой (самоопылении у растений) расщепления по изучаемым признакам не наблюдалось.
Полученные данные Мендель обработал математически, в результате чего получилась четкая закономерность наследования отдельных признаков родительских форм их потомками в ряде последующих поколений. Эту закономерность Мендель сформулировал в виде правил наследственности, получивших позднее название законов Менделя.
Первый закон Менделя — закон единообразие гибридов первого поколения (правило доминирования): при скрещивании гомозиготных особей, отличающихся одной или несколькими парами альтернативных признаков, все гибриды первого поколения окажутся по этим признакам единообразными. У гибридов проявятся доминантные признаки одного из родителей
Г. Мендель изучал наследование цвета семян гороха (его альтернативные варианты — желтый или зеленый — кодируются парой аллелей одного гена):
|
Р (родители) |
АА Х аа |
|
|
G (гаметы) |
А |
а |
|
F1 (гибриды первого поколения) |
Аа |
|
Гибриды первого поколения оказались единообразными по исследуемому признаку. В F| проявился лишь один (желтый) из пары альтернативных вариантов признака цвета семян, названный доминантным.
Из гибридных семян гороха Г. Мендель вырастил растения, которые в результате самоопыления произвели семена второго поколения, как с желтой, так и с зеленой окраской семядолей. У части гибридов F2 вновь возник признак, не обнаруженный у гибридов F1. Этот признак (зеленый) назван рецессивным. Соотношение потомков с доминантным и рецессивным проявлением признака оказалось близко к 3/4 с доминантным признаком (желтые семена) и 1/4 с рецессивным признаком (зеленые семена).
|
Р |
Аа Х Аа |
|
|
G |
А, а |
А, а |
|
F1 |
1/4АА, 1/4Аа, 1/4Аа, 1/4аа |
|
Второй закон Менделя можно сформулировать следующим образом: при моногибридном скрещивании гетерозиготных особей (гибридов F1) во втором поколении наблюдается расщепление по вариантам анализируемого признака в отношении 3:1 по фенотипу и 1:2:1 по генотипу.
Цитологические основы наследования. Закон “чистоты гамет”
Для объяснения распределение признаков у гибридов последовательных поколений Г. Мендель предположил, что каждый наследственный признак зависит от наличия в соматических клетках двух наследственных факторов, полученных от отца и матери. К настоящему времени установлено, что наследственные факторы Менделя соответствую г генам, расположенным в хромосомах, полому сходство между поведением наследственных факторов и поведением хромосом при мейозе и оплодотворении стало очевидным.
Гомозиготные растения с желтыми семенами имеют генотип АА и образуют гаметы одного сорта с аллелью А; растения с зелеными семенами имеют генотип аа и образуют гаметы с аллелью а. В момент оплодотворения происходит слияние половых клеток и возникают гетерозиготные диплоидные особи с генотипом Аа, образующие семена с доминантной желтой окраской.
В F1 во время анафазы первого деления мейоза гомологичные хромосомы с аллелями А и а отходят к разным полюсам клетки и затем попадают в разные гаметы, причем яйцеклеток с аллелем А и с аллелем а образуется примерно в равном количестве, также как и спермиев с А и а. Вероятность оплодотворения яйцеклеток с А и а аллелями спермиями с А аллелем равна вероятности оплодотворения их спермиями с а аллелем.
Соотношение генотипов особей гибридных поколений можно показать графически при помощи решетки Пеннета. При ее составлении гаметы одного из родителей выписывают снаружи решетки по вертикали, гаметы другого — по горизонтали. Возникающие в результате оплодотворения зиготы вписываются внутри в клетках на пересечении линий, идущих от соответствующих гамет.
|
Гаметы женского организма |
|||
|
1/2А |
1/2а |
||
|
Гаметы мужского организма |
1/2А |
1/4АА |
1/4Аа |
|
1/2а |
1/4Аа |
1/4аа |
|
Таким образом, правило чистоты гамет можно сформулировать следующим образом: в процесс образования половых клеток в каждую гамету попадает только один ген из аллельной пары.
Дигибридное скрещивание. Третий закон Менделя (закон независимого комбинирования)
Дигибридным называют скрещивание, при котором прослеживается наследование по двум парам альтернативных признаков. В опытах Менделя при скрещивании гомозиготного сорта гороха, имевшего желтые АА и гладкие ВВ семена, с сортом гороха с зелеными аа и морщинистыми bb семенами, гибриды F1 имели желтые и гладкие семена, т.е. проявились доминантные признаки (гибриды единообразны).
|
Р |
ААВВ Х ааbb |
|
|
G |
АB |
аb |
|
F1 |
АаBb |
|
При скрещивании дигетерозигот (особей F1) во втором поколении гибридов (F2) будет наблюдаться расщепление признаков по фенотипу в соотношении 9:3:3:1.
|
Р |
АаВb |
X |
АаВb |
|
|
Гаметы |
1/4АB |
1/4Аb |
1/4аВ |
1/4аb |
|
1/4АB |
1/16АABB |
1/16АABb |
1/16АaBB |
1/16АaBb |
|
1/4Аb |
1/16АABb |
1/16АAbb |
1/16АaBb |
1/16Аabb |
|
1/4аВ |
1/16АaBB |
1/16АaBb |
1/16aaBB |
1/16aaBb |
|
1/4аb |
1/16АaBb |
1/16Аabb |
1/16aaBb |
1/16aabb |
Гибридные семена в F2 распределились на четыре фенотипические группы в соотношении: 9/16 — с гладкими желтыми (А-В-), 3/16 — с гладкими зелеными (A-bb), 3/16 — с морщинистыми желтыми (ааВ-) и 1/16 — с зелеными морщинистыми семенами (aabb). Таким образом, форма семян комбинировалась независимо от цвета семян и у потомков F2 наблюдались новые комбинации признаков. Это явление получило название закона независимого комбинирования: гены разных а цельных пар и соответствующие им признаки наследуются потомками независимо друг от друга, комбинируясь в различных сочетаниях.
Одна пара аллельных генов комбинируется независимо от другой пары аллельных генов, так как они расположены в разных парах гомологичных хромосом.
При образовании гамет дигетерозигота АаВb образует четыре типа гамет. В анафазе мейоза I гомологичные хромосомы каждой пары расходятся к разным полюсам клетки независимо от других пар гомологичных хромосом, поэтому аллели одной пары распределяются по гаметам независимо oт аллелей других пар. Каждая гамета может получить только один ген из каждой аллельной пары. Таким образом, каждая дигетерозигота образует четыре сорта гамет примерно в одинаковом количественном соотношении. Соответственно независимому распределению аллельных генов каждой пары гомологичных хромосом осуществляется и независимое наследование обусловленных ими признаков. Любая женская гамета имеет равные шансы быть оплодотворенной любой мужской.
Расщепления, полученные Г. Менделем, носят статистический характер. т.е. определяются вероятностным характером комбинирования хромосом в мейозе и гамет при оплодотворении. В связи с этим экспериментальные данные не соответствуют абсолютно точно теоретически ожидаемым результатам. Только специальные методы статистической обработки позволяют установить достоверность соответствия практических результатов теоретически ожидаемым.
Анализирующее скрещивание
При полном доминировании гетерозиготы (Аа) и доминантные гомозиготы (АА) характеризуются одинаковым доминантным фенотипом. Их генотипы можно различить только с помощью гибридологического анализа, т.е. по потомству, которое образуется в результате скрещивания, получившего название анализирующего. Анализирующим является такой тип скрещивания, при котором испытуемую особь с доминантным признаком скрещивают с особью, гомозиготной по рецессивному аллелю.
Если доминантная особь гомозиготна, потомство от такого скрещивания будет единообразным и расщепления не произойдет. В том случае, если особь с доминантным признаком гетерозиготна, расщепление произойдет в отношении 1:1 по фенотипу и генотипу при моногибридном скрещивании.
|
P Aa X aa |
|
|
G A, a |
a |
|
F1 Aа, аа |
|
При дигибридном анализирующем скрещивании расщепление по фенотипу и генотипу составит 1:1:1:1.
|
Р |
АаВb |
X |
aabb |
|
|
1/4AВ |
1/4Ab |
1/4аВ |
1/4ab |
|
|
ab |
1/4АаВb |
1/4Aabb |
1/4ааВb |
1/4ааbb |
Такой результат скрещивания прямо показывает, что у гетерозиготного организма образуется четыре сорта гамет, причем в приблизительно равном соотношении (по 25%). Одинаковые же гаметы гомозиготного рецессива как бы анализируют генотип организма, имеющего доминантный фенотип.
Сцепленное наследование. Хромосомная теория наследственности
Число генов у каждого организма значительно больше числа хромосом. Например, в геноме человека идентифицировано 20—25 тыс. генов. Следовательно, в каждой хромосоме находится много генов. Работы Т. Моргана и его учеников в начале 20-х гг. XX в., показали, что гены, локализованные в одной хромосоме, наследуются вместе или сцеплено, т.е. передаются в поколениях, преимущественно не обнаруживая независимого наследования. Гены, расположенные в одной хромосоме, называют группой сцепления. Так как гомологичные хромосомы содержат аллельные гены, то в группу спепления включают обе гомологичные хромосомы, поэтому число групп спепления соответствует гаплоидному набору. У человека 23 группы сцепления, у плодовой мушки (дрозофилы) — 4, у гороха — 7 и т.д. Гомологичные хромосомы могут обмениваться аллельными генами в процессе кроссинговера (профаза первого деления мейоза), частота которого зависит от расстояния между генами.
Гены могут быть расположены в хромосоме непосредственно друг за другом. В таком случае кроссинговер между ними практически невозможен. Они почти всегда наследуются вместе. Такое спепление называют полным. Т. Морган изучал наследование двух признаков (цвет тела и длина крыльев) у дрозофилы. При скрещивании гетерозиготного самца (BbVv) с серым телом и нормальной длиной крыльев с гомозиготной рецессивной самкой (bbvv — черное тело и короткие крылья) образуется всего два фенотипических класса потомков: 50% мух с серым телом и длинными крыльями и 50% мух с черным телом и короткими крыльями. Комбинаций признаков у потомков не происходило (рис. 7.1).

Рис. 7.1. Полное сцепление
У самцов дрозофилы кроссинговер практически не происходит, поэтому гены у дигетерозиготных самцов, расположенные в одной хромосоме, обнаруживают полное сцепление, т.е. наследуются вместе.
Если в анализирующем скрещивании поменять местами родительские формы, т.е. скрестить дигибридную самку с гомозиготным рецессивным самцом (рис. 7.2), то образуются четыре категории потомков: 41,5% мух серых с длинными крыльями, 41,5% — черных с укороченными крыльями, 8,5% — серых с укороченными крыльями и 8,5% — черных с длинными крыльями. Если бы каждая пара генов находилась в разных парах гомологичных хромосом (независимое наследование), то число сортов потомков должно быть примерно одинаковым (по 25% каждого из четырех сортов; см. параграф “Анализирующее скрещивание”). Такое отклонение от ожидаемого при независимом расщеплении свидетельствует о наличии сцепления. Однако в отличие от скрещивания, где гетерозиготным был самец, у гетерозиготных самок в мейозе возможен обмен (кроссинговер) между локусами гомологичных хромосом, где находятся гены В и V. Рекомбинантные типы потомства при таком скрещивании возникают из кроссоверных гамет, которые несут рекомбинантные хромосомы, образующиеся у самки в процессе кроссинговера при мейозе. Такое сцепление генов называют неполным.

Рис. 7.2. Неполное сцепление
Расстояние между генами выражают в процентах кроссинговера между ними. Процент кроссинговера между двумя генами равен сумме процентов особей с новыми (отличными от родителей) комбинациями признаков. Так, в рассмотренном выше примере 17% потомков было с новыми комбинациями признаков (8,5% + 8,5%). Следовательно, расстояние между генами В и V равно 17%.
Частота кроссинговера зависит от расстояния между генами одной группы сцепления. Чем дальше гены расположены в хромосоме, тем кроссинговер между ними чаше и наоборот. Следовательно, о расстоянии между генами в хромосоме можно судить по частоте кроссинговера. Т. Морган и его сотрудники показали, что, установив частоту рекомбинаций (процента кроссинговера) между генами, относящимися к одной группе сцепления, можно построить карты хромосом с нанесенным на них порядком расположения генов. Например, если известно, что расстояние между двумя генами одной группы сцепления А и В составило 4,6% и требуется установить место расположения третьего гена С этой же хромосомы, то достаточно выяснить процент кроссинговера между геном С и генами А и В. Если процент кроссинговера межу генами А и С составил 3,3%, а между генами В и С — 1,3%, то ген С расположен между генами А и В. Если же процент кроссинговера между генами А и С составил 5,7%, то ген С расположен на одном из концов хромосомы.
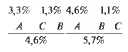
Хромосомное определение пола. Сцепленное с полом наследование
При изучении кариотипов клеток многих видов животных и растений было установлено, что в ядрах соматических клеток содержится двойной набор аутосом, одинаковый у самцов и самок, и две половые хромосомы, по которым различаются оба пола. Так, например, у человека в соматических клетках женского организма имеется 44 аутосомы и две половые хромосомы — XX. У мужчин имеются также 44 аутосомы и одна X и одна Y-хромосома (рис. 7.3). Y-хромосома у млекопитающих, большинства насекомых, многих рыб, растений и у других организмов является определяющей в развитии мужского пола. При созревании половых клеток в результате мейоза гаметы получают гаплоидный набор хромосом. Все яйцеклетки имеют 22 аутосомы и Х-хромосому. Сперматозоиды будут двух сортов. 50% при сперматогенезе получат 22 аутосомы и Y-хромосому, другая половина — 22 аутосомы и Х-хромосому. Пол будущего организма будет определяться при оплодотворении:
• если в яйцеклетку попадет Y-содержащий сперматозоид, то из зиготы разовьется особь мужского пола;
• если X-содержащий — то особь женского пола.
Таким способом поддерживается первичное соотношение полов 1:1, что является прямым результатом хромосомного механизма определения пола. Женский пол, который образует гаметы, одинаковые по половой хромосоме, называют гомогаметным, а мужской пол, образующий разные гаметы по X и Y-хромосомам, — гетерогаметным. У млекопитающих (в том числе человека), червей, ракообразных, большинства насекомых (в том числе дрозофилы), большинства земноводных, некоторых рыб гомогаметным является женский пол, гетерогаметным — мужской.

Рис. 7.3. Хромосомный набор (кариотип) человека:
а — мужчины, б — женщины
У некоторых насекомых (например, водяного клопа, кузнечика и др.) Y-хромосома вообще отсутствует (рис. 7.4). В этих случаях у самцов имеется всего одна Х-хромосома. В результате половина сперматозоидов имеет половую хромосому, а другая — ее лишена.
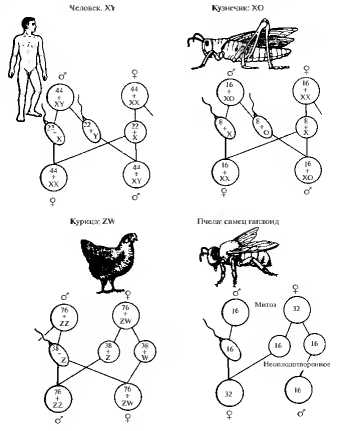
Рис. 7.4. Основные типы хромосомного определения
У птиц, пресмыкающихся, некоторых земноводных и рыб, части насекомых (бабочки и ручейники) гетерогаметным является женский пол, а мужской гомогаметным. В этом случае для обозначения половых хромосом используют другие символы. Например, у кур, имеющих в соматических клетках 78 хромосом, хромосомная формула мужского пола 76А + ZZ, женского - 76А + ZW.
У пчел и муравьев нет половых хромосом: самки диплоидны, самцы гаплоидны. Самки развиваются из оплодотворенных яйцеклеток, трутни — из неоплодотворенных.
Сцепленное с полом — наследуются признаки, гены которых находятся в половых хромосомах. У многих видов X- и Y-хромосомы резко различны по величине. Как правило, Y-хромосома невелика по размерам и не содержит аллелей многих генов, расположенных в Х-хромосоме (рис. 7.5). Таким образом, у мужчин большинство генов, локализованных в Х-хромосоме, находятся в гемизиготном состоянии, т.е. представлены в единственном числе и не имеют аллельной пары. Наследование, сцепленное с X-хромосомой, может быть доминантным и рецессивным. Например, у человека с Х-хромосомой сцепленно наследуются рецессивные болезни дальтонизм (цветовая слепота) и гемофилия (несвертываемость крови). Они встречаются чаще у мужчин.
|
р |
XHXh X XHY |
|
|
G |
ХH, Xh |
ХH, Y |
|
F1 |
XHXH, XHY, XHXh, XhY |
|
Н — нормальная свертываемость, h — гемофилия
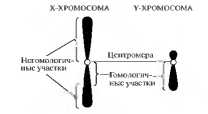
Рис. 7.5. X- и Y-хромосомы
В брак вступили здоровый мужчина и женщина — носительница рецессивного гена гемофилии. Вероятность рождения больного мальчика в этой семье составит 50%. Девочки — здоровы, но половина будет носителями гена гемофилии.
При локализации гена в Y-хромосоме признаки передаются от отца к сыну. В Y-хромосоме находится ген, обусловливающий дифференцировку мужского пола и ряд других генов.
Цитоплазматическая наследственность
Помимо нуклеиновых кислот, входящих в состав хромосом и обусловливающих хромосомное наследование, в цитоплазме клеткок про- и эукариот находятся молекулы ДНК (реже РНК), реплицирующиеся либо в виде автономных структур, либо в составе органоидов. Количество нехромосомной ДНК сравнительно невелико и составляет для разных организмов от десятых долей до нескольких процентов. Нехромосомные молекулы нуклеиновых кислот, реплицирующиеся автономно от хромосом в клетках эукариот и бактерий, называют общим термином — плазмиды. В составе плазмид могут находиться гены, кодирующие ряд признаков клеток-хозяев. Наследование этих генов не подчинено менделевским закономерностям наследования признаков.
У растений было обнаружено наследование пестролистности. Признак пестролистности связан с мутациями в ДНК хлоропластов, нарушающими синтез хлорофилла. Вследствие этого отдельные части листа и других зеленых органов растений лишены хлорофилла и оказываются светлыми. Во время мейоза хлоропласты попадают в цитоплазму яйцеклеток, а в клетках пыльцы большинства видов растений они практически отсутствуют. Таким образом, наследование пестролистности передается по материнской линии.
Митохондрии также содержат кольцевидные молекулы ДНК и имеют собственный аппарат белкового синтеза. Митохондриальные гены кодируют в основном две группы признаков, связанных с работой ферментативных систем синтеза АТФ и устойчивостью к антибиотикам и другим клеточным ядам. У многоклеточных организмов, размножающихся половым путем, передача в поколениях признаков, обусловленных митохондриальными генами, возможна только от материнского организма. Это связана с тем, что яйцеклетка в цитоплазме содержит много митохондрий, а сперматозоид практически лишен цитоплазмы.
Взаимодействие генов
Генотип любого организма представляет собой сложную систему взаимодействующих генов, как аллельных, так и неаллельных. Развитие признаков является результатом взаимодействия генов, точнее, взаимодействия не самих генов (участков молекул ДНК), а образуемых на основе их генетической информации генных продуктов (РНК, а затем белков). Синтезируемые в клетках организма белки, образуя структуры или управляя процессами обмена веществ, играют важную роль в процессах формирования фенотипа организма.
При взаимодействии аллельных генов возможны разные варианты проявления признака. Если аллели находятся в гомозиготном состоянии, то развивается соответствующий аллелю вариант признака. В случае гетерозиготности развитие признака будет зависеть от конкретного вида взаимодействия аллельных генов.
При полном доминировании фенотип гетерозиготы (Аа) не отличается от фенотипа доминантной гомозиготы (АА). Рецессивный признак имеют только рецессивные гомозиготы (аа). Примером полного доминирования может служить высокий рост растений над карликовостью, вьющиеся волосы над прямыми у человека и т.д.
Неполное доминирование наблюдается в случаях, когда фенотип гетерозигот (Аа) отличается от фенотипа гомозигот (АА и аа) промежуточной степенью проявления признака. Аллель, отвечающий за формирование признака, находясь в двойной дозе у гомозиготы (АА), проявляется интенсивнее, чем в одинарной дозе у гетерозиготы (Аа). Примером расщепления при неполном доминировании может служить наследование окраски цветков Ночной красавицы.
|
Р АА (растения с красными цветками) X аа (растения с белыми цветками) |
|
|
С А |
а |
|
F1 |
Аа (растения с розовыми цветками) |
|
Р |
Аа |
Х |
Аа |
|
|
G |
А, а |
А, а |
||
|
F2 |
АА |
Аа |
Аа |
аа |
В F2 наблюдается расщепления 1:2:1 как по фенотипу, так и по генотипу.
При кодоминировании у гетерозиготных организмов каждый из аллельных генов вызывает формирование в фенотипе контролируемого им признака (антигена А и антигена В). Примером этой формы взаимодействия аллелей служит развитие IV группы крови человека по системе АВО.
|
Р |
IAIA |
X |
IBIB |
|
G |
IA |
IB |
|
|
F1 |
IAIB |
||
Если человек гетерозиготен IAIB, его эритроциты имеют два поверхностных антигена: А В (IV группа).
Неаллельные гены — это гены, расположенные в негомологичных участках хромосом и кодирующие неодинаковые белки. Неаллельные гены также могут взаимодействовать между собой. Это приводит к значительным численным отклонениям расщеплений по фенотипу от установленных Менделем при дигибридном скрещивании. Различают три основных типа взаимодействия неаллельных генов: комплементарносгь, эпистаз и полимерию.
Под комплементарностью понимают такой тип взаимодействия двух доминантных неаллельных генов, при котором эти гены вместе обусловливают развитие нового признака, отличного от родительских вариантов. Так, у душистого горошка есть ген А, обусловливающий синтез бесцветного предшественника пигмента - пропигмента. Ген В определяет синтез фермента, под действием которого из пропигмента образуется пигмент. Цветки душистого горошка с генотипом ааВВ и ААbb имеют белый цвет: в первом случае есть фермент, но нет пропигмента, во втором — есть пропигмент, но нет фермента, переводящего пропигмент в пигмент. При скрещивании двух растений душистого горошка с белыми цветками (ААbb x ааВВ) получается гибрид с пурпурной окраской цветков (АаВb), а в F2 будет наблюдаться расщепление: 9/16 растений будут с пурпурными лепестками цветов и 7/16 с белыми (9:7).
|
Р |
АаВb |
X |
АаВb |
|
|
Гаметы |
1/4АB |
1/4Аb |
1/4аВ |
1/4аb |
|
1/4АB |
1/16АABB |
1/16АABb |
1/16АaBB |
1/16АaBb |
|
1/4Аb |
1/16АABb |
1/16АAbb |
1/16АaBb |
1/16Аabb |
|
1/4аВ |
1/16АaBB |
1/16АaBb |
1/16aaBB |
1/16aaBb |
|
1/4аb |
1/16АaBb |
1/16Аabb |
1/16aaBb |
1/16aabb |
Лепестки душистого горошка с генотипами ааВВ, aaBb, ААаа, Aabb, aabb имеют белый цвет. Во всех остальных генотипах присутствуют оба доминантных неаллельных гена, что обусловливает образование пропигмента и фермента, участвующего в образовании пурпурного пигмента.
Эпистаз — взаимодействие неаллельных генов, при котором один из них подавляется другим. Подавляющий ген называется эпистатичным, подавляемый — гипостатичным. Эпистатическое взаимодействие неаллельных генов может быть доминантным и рецессивным.
Доминантный эпистаз наблюдается при наследовании окраски оперения кур. Ген С в доминантной форме определяет нормальную продукцию пигмента, доминантный ген I является энис готическим по отношению к гену С (I > С). В результате этого куры, имеющие в генотипе доминантный аллель гена окраски, в присутствии эпистатического гена оказываются белыми. При скрещивании двух белых птиц (IICC и iicc) все особи F1 тоже будут белыми (IiСс), но в F2 произойдет расщепление по фенотипу в отношении: 13/16 — белых и 3/16— окрашенных. Особи с генотипами iicc и I-С- (вторые аллели могут быть как доминантными, так и рецессивными) будут белыми, a iiC — окрашенными.
|
р |
liCc |
X |
liCc |
|
|
Гаметы |
1/4IC |
1/4Ic |
1/4iC |
1/4ic |
|
1/4IC |
1/16IICC |
1/16IICc |
1/16IiCC |
1/16IiCc |
|
1/4Iс |
1/16IICc |
1/16IIcc |
1/16IiCc |
1/16Iicc |
|
1/4iС |
1/16IiCC |
1/16IiCc |
1/16iiCC |
1/16iiCc |
|
1/4ic |
1/16IiCc |
1/16iiCc |
1/16iiCc |
1/16iiicc |
Рецессивный эпистаз характерен при наследовании окраски шерсти у домовых мышей. Окраска агути (рыжевато-серая окраска шерсти) определяется доминантным геном А. Его рецессивный аллель а в гомозиготном состоянии обусловливает черную окраску. Доминантный ген другой пары В не препятствует синтезу пигмента, гомозиготы по рецессивному аллелю bb являются альбиносами с белой шерстью и красными глазами (отсутствие пигмента в шерсти и радужной оболочке глаз). При скрещивании черной мыши с генотипом aabb и белой — с генотипом ААbb все потомки в F1 (АаВb) будут иметь окраску агути. В F2 расщепление по фенотипу происходит следующим образом: 9/16— агути, 3/16 — черных и 4/16— белых. Такое расщепление обусловлено гомозиготностью по эпистатическому гену bb.
|
Р |
Аа |
X |
АаВb |
|
|
Гаметы |
1/4АB |
1/4Аb |
1/4аВ |
1/4аb |
|
1/4АB |
1/16АABB |
1/16АABb |
1/16АaBB |
1/16АaBb |
|
1/4Аb |
1/16АABb |
1/16АAbb |
1/16АaBb |
1/16Аabb |
|
1/4аВ |
1/16АaBB |
1/16АaBb |
1/16aaBB |
1/16aaBb |
|
1/4аb |
1/16АaBb |
1/16Аabb |
1/16aaBb |
1/16aabb |
Полимерия — такое взаимодействие доминантных неалельных генов (полигенов), в результате которого степень развития признака зависит от числа этих генов в генотипе организма. Чем больше в генотипе доминантных неаллельных генов, тем сильнее выражен тот или иной признак. Признаки, зависящие от полимерных генов, относят к количественным признакам (рост, вес у животных, длина колоса у злаков, количество сахара в корнеплодах сахарной свеклы и т.п.). Принципиальной особенностью количественных признаков является то, что различия по ним между отдельными организмами могут быть очень небольшими и требуют точных измерений; в отличие от качественных альтернативных признаков разница между некоторыми из них велика и видна просто при наблюдении.
Рассмотрим в качестве примера наследование цвета кожи у человека. Допустим, что темный цвет кожи зависит от двух пар доминантных неаллельных генов (А1А1А2А2). Рецессивные аллели этих генов обусловливают светлый цвет кожи человека (а1а1а2а2). Дети от брака мужчины негроидной расы и белой женщины будут мулатами.
|
Р |
А1А1А2А2 |
X |
а1а1а2а2 |
|
G |
А1А2 |
а1а2 |
|
|
F1 |
А1а1А2а2 |
В потомстве от двух мулатов вероятность рождения негра (А|А,А2А2) или белового (а|а]а2а2) будет равна '/|б. Остальные генотипы попадают в промежуточные фенотипические классы.
|
Р |
A1a1A2a2 |
X |
A1a1A2a2 |
|
|
Гаметы |
1/4А1А2 |
1/4А1а2 |
1/4а1А2 |
1/4а1а2 |
|
1/4А1А2 |
1/16А1А1А2А2 |
1/16А1А1А2А2 |
1/16А1а1А2А2 |
1/16А1а1А2а2 |
|
1/4А1а2 |
1/16А1А1А2а2 |
1/16А1А1а2а2 |
1/16А1а1А2а2 |
1/16А1а1а2а2 |
|
1/4а1А2 |
1/16А1а1А2А2 |
1/16А1а1А2а2 |
1/16а1а1А2А2 |
1/16а1а1А2а2 |
|
1/4а1а2 |
1/16А1а1А2а2 |
1/16А1а1а2а2 |
1/16а1а1А2а2 |
1/16а1а1а2а2 |
Чем больше доминантных генов в генотипе, тем темнее кожа, и наоборот.
Множественное (плейотропное) действие гена. Один и тот же ген может влиять на формирование ряда признаков организма. Например, ген, вызывающий образование бурой семенной кожуры у гороха, способствует развитию пигмента и в других частях растения. У человека есть ген, определяющий рыжую окраску волос. Этот же ген обусловливает более светлую окраску кожи, а также появление веснушек. Для большинства генов с той или иной степенью плейотропии характерно сильное влияние на один признак и значительно более слабое влияние на другие. На уровне первичного действия ген имеет единственную функцию — кодирование одного полипептида. Плейотропный эффект проявляется в том, что действие фермента помимо влияния на один признак отражается на вторичных реакциях биосинтеза которые в свою очередь влияют на формирование различных признаков организма. Действие гена может быть изменено соседством других генов и условиями внешней среды. Таким образом, в онтогенезе действуют не отдельные гены, а весь генотип как целостная сбалансированная система со сложными связями и взаимодействиями между ее компонентами. Эта система динамична: появление в результате мутаций новых аллелей или генов, формирование новых хромосом и даже новых геномов приводят к заметному изменению генотипа во времени.
Рассмотрев различные виды взаимодействия генов, можно считать, что генотип является сбалансированной системой взаимодействующих генов, развитие признака есть результат проявления нескольких генов в конкретных условиях среды.