БИОЛОГИЯ - справочник для старшеклассников и поступающих в вузы
Закономерности изменчивости - ОСНОВЫ ГЕНЕТИКИ И СЕЛЕКЦИИ - ОБЩАЯ БИОЛОГИЯ
Роль генотипа и условий внешней среды в формировании фенотипа. Модификационная изменчивость. Норма реакции. Статистические закономерности модификационной изменчивости. Мутации, их причины. Мутационная изменчивость. Закон гомологических рядов наследственной изменчивости Н. И. Вавилова. Экспериментальное получение мутаций.
Мутации как материал для искусственного и естественного отбора. Загрязнение природной среды мутагенами и его последствия. Генетика и теория эволюции. Генетика популяций. Формы естественного отбора: движущий и стабилизирующий.
Задание 12
• Повторить имеющийся по теме материал.
• Ответить на вопросы для самоконтроля.
• Выполнить контрольную работу № 20.
• Проанализировать схемы 22-24, таблицу 51.
• Проверить по словарю знание терминов.
Вопросы для самоконтроля
• Что такое ген?
• Что такое генотип?
• Какие гены называют аллельными?
• От чего зависит фенотип?
• Какие признаки фенотипа пластичны?
• Чем ограничена пластичность признаков?
• Как называется изменчивость фенотипа?
• Как называется изменчивость генотипа?
• Какие структуры клетки перестраиваются при мутационной изменчивости?
• Под воздействием каких факторов происходят генные мутации?
• В результате какого явления осуществляются хромосомные мутации?
• Что такое полиплоидия?
• Что более подвержено изменению при неблагоприятных условиях окружающей среды — фенотип или генотип?
• Что называют нормой реакции?
• Как объяснить предвидение Н. И. Вавилова о существовании в природе безалкалоидного (неядовитого) люпина?
• Что принято в современной теории эволюции за элементарную эволюционную единицу?
• Какова роль мутаций в природных популяциях?
• Почему теорию эволюции нашего времени называют синтетической и кто ее создал?
Контрольная работа № 20
1. Что характерно для мутации (возникает при скрещивании, при кроссинговере, возникает внезапно в ДНК или в хромосомах)?
2. Признаки какой изменчивости передаются потомству (модификационной, мутационной)?
3. Что подвергается изменениям при возникновении мутаций (генотип, фенотип)?
4. Наследуются признаки генотипа или фенотипа?
5. Для какой изменчивости характерны следующие признаки: возникают внезапно, могут быть доминантными или рецессивными, полезными или вредными, наследуются, повторяются (мутационная, модификационная)?
6. Где происходят мутации (в хромосомах, в молекулах ДНК, в одной паре нуклеотидов, в нескольких нуклеотидах)?
7. В каком случае мутация проявляется фенотипически (в любом, в гомозиготном организме, в гетерозиготном организме)?
8. Какова роль мутаций в эволюционном процессе (увеличение изменчивости, приспособление к окружающей среде, самосовершенствование организма)?
9. От чего зависит фенотип (от генотипа, от окружающей среды, ни от чего не зависит)?
10. Чем определяется размах изменчивости признаков организма (окружающей средой, генотипом)?
11. Признаки какой изменчивости выражаются в виде вариационного ряда и вариационной кривой (мутационной, модификационной)?
12. Какие признаки обладают узкой нормой реакции (качественные, количественные), какие более пластичны (качественные, количественные)?
13. Какая форма естественного отбора в популяции приводит к образованию новых видов (движущий, стабилизирующий), какая — к сохранению видовых признаков (движущий, стабилизирующий)?
Схема 22. Формы изменчивости
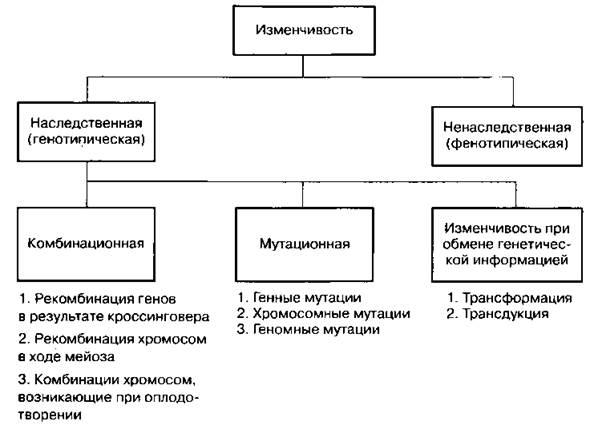
Схема 23. Классификация мутаций

Схема 24. Классификация мутаций по уровню их возникновения
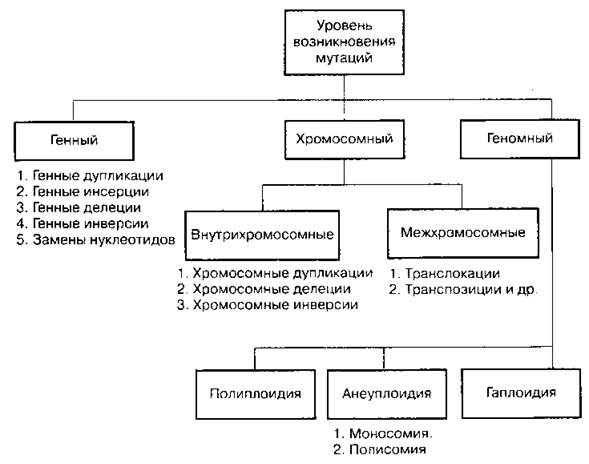
Таблица 51. Сравнительная характеристика форм изменчивости
|
Характеристика |
Модификационная изменчивость |
Мутационная изменчивость |
|
Объект изменения |
Фенотип в пределах нормы реакции |
Генотип |
|
Отбирающий фактор |
Изменение условий окружающей среды |
Изменение условий окружающей среды |
|
Наследование признаков |
Не наследуются |
Наследуются |
|
Подверженность изменениям хромосом |
Не подвергаются |
Подвергаются при хромосомной мутации |
|
Подверженность изменениям молекул ДНК |
Не подвергаются |
Подвергаются в случае генной мутации |
|
Значение для особи |
Повышает или понижает жизнеспособность, продуктивность, адаптацию |
Полезные изменения приводят к победе в борьбе за существование, вредные — к гибели |
|
Значение для вида |
Способствует выживанию |
Приводит к образованию новых популяций, видов и т. д. в результате дивергенции |
|
Роль в эволюции |
Приспособление организмов к условиям среды |
Материал для естественного отбора |
|
Форма изменчивости |
Определенная (групповая) |
Неопределенная (индивидуальная) |
|
Подчиненность закономерности |
Статистическая закономерность вариационных рядов |
Закон гомологических рядов наследственной изменчивости |
Наследование некоторых признаков, сцепленных с полом, у человека
У человека в Х-хромосоме помимо генов, участвующих в развитии половых признаков, находятся гены, определяющие свертываемость крови и цветоразличение, при этом доминантный аллель определяет нормальное формирование признака, а рецессивный — патологию (гемофилию и дальтонизм соответственно). Рассмотрим наследование этих генов на примере наследования дальтонизма (ген гемофилии наследуется аналогично). Ген дальтонизма является универсально распространенным на нашей планете. Фенотипически он проявляется у 7% мужчин и у 0,5% женщин, но носительницами этого гена являются 13% женщин (т. е. они имеют этот ген в гетерозиготном состоянии). Итак, у человека в Х-хромосоме находится ген, определяющий восприятие красного и зеленого цветов. Его доминантный аллель D обусловливает нормальное цветоразличение, в то время как рецессивный аллель d вызывает дальтонизм (неспособность различать красный и зеленый цвета). Поскольку ген локализован в Х-хромосоме, то в гетерозиготном состоянии он фенотипически не проявляется и женщина-носительница имеет нормальное цветовое различение* Рассмотрим наследование этого признака на различных примерах.
1. Брак женщины — носительницы гена дальтонизма (XDXd) и здорового мужчины (XDY)
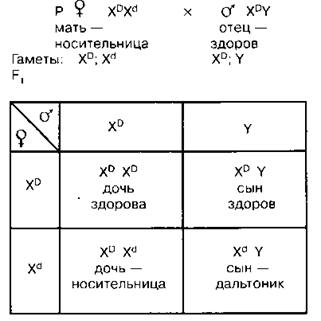
Половина дочерей от этого брака будут здоровы, а половина будут носительницами; половина сыновей будут здоровы, а половина — дальтониками.
2. Брак здоровой женщины и мужчины-дальтоника

Обратите внимание на то, что в этом браке все сыновья отца-дальтоника будут здоровы (поскольку от отца они получат не несущую данного гена Y-хромосому), а все дочери будут носительницами (поскольку от отца-дальтоника они получат свою вторую Х-хромосому).
3. Брак женщины-носительницы и мужчины-дальтоника
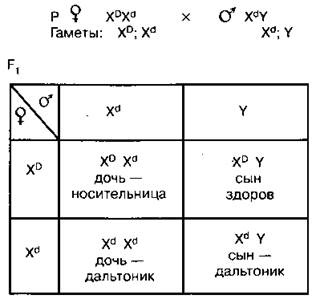
От этого брака половина дочерей будут носительницами, а половина — дальтониками; половина сыновей будут дальтониками, а половина — здоровы. Этот пример наглядно иллюстрирует опасность браков между близкими родственниками, так как в этом случае велика вероятность перехода рецессивных генов, вызывающих различные патологии, в гомозиготное состояние.
4. Брак женщины-дальтоника и здорового мужчины
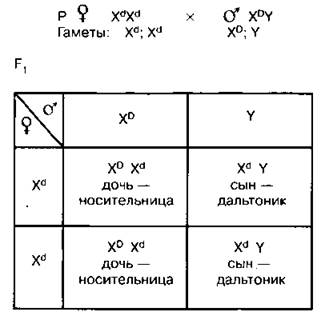
Обратите внимание, что в данном браке все сыновья здорового мужчины будут дальтониками (поскольку свою Х-хромосому они получат от матери, а у нее обе хромосомы несут ген дальтонизма); все дочери будут носительницами.
Опыты Моргана, позволившие ему сформулировать хромосомную теорию наследственности
Перед тем как начать разговор о создании Т. Морганом в 1911 г. хромосомной теории наследственности, давайте составим представление о том, какие сведения уже были накоплены биологией к этому времени.
Исследователи конца XIX в. считали, что во время деления клетки ядро полностью исчезает и в новых клетках ядра всегда возникают заново. Однако к началу XX в. наука накопила сведения, полностью опровергнувшие эти взгляды. Были открыты митоз (1874 г. — у растительных клеток; 1878 г. — у животных) и мейоз (1882 г. — у животных; 1888 г. — у растений). Исследования этих процессов показали, что ядро не исчезает в ходе клеточного деления и не возникает затем вновь в дочерних клетках. Стало очевидным, что между ядром материнской и дочерней клетки существует преемственность, которая заключается в равномерном распределении хромосом между клетками в ходе клеточных делений.
В 1900 г. были вновь открыты законы генетики (К. Корренс, Э. Чермак, Г. де Фриз), а в 1902-1903 гг. У. Саттон (США) и Т. Бовери (Германия) независимо друг от друга предположили, что наследственные задатки (гены) расположены в хромосомах. (Термин «ген» был предложен позже — в 1909 г. В. Иогансеном.) Параллелизм в поведении генов (при скрещиваниях и в расщеплении потомства) и хромосом (в процессе образования гамет) говорил в пользу того, что гены расположены именно в хромосомах.
В 1906 г. Р. Пеннет и У. Бетсон впервые описали нарушение менделевского закона независимого наследования двух признаков. В их опытах на душистом горошке два признака — окраска цветков и форма пыльцы — не давали независимого расщепления в потомстве, и потомки оказывались похожи на родителей. Постепенно таких исключений, не подчинявшихся закону независимого наследования признаков, накапливалось все больше и больше, и поэтому все более актуальными становились поиски ответов на следующие вопросы: почему далеко не все признаки наследуются независимо друг от друга и как именно такие признаки наследуются? Как именно расположены гены в хромосомах? (Ведь любой организм несет очень много признаков, а число его хромосом невелико.) Каковы закономерности наследования генов, находящихся в одной хромосоме? На эти вопросы смогла ответить хромосомная теория наследственности, сформулированная в 1911 г. Т. Морганом. К своим выводам он пришел, изучая наследование различных признаков у плодовой мухи дрозофилы. Обратимся и мы к результатам его экспериментов.
У мухи дрозофилы ген серого цвета тела (В) доминирует над геном черного цвета (b), а ген нормальной длины крыльев (V) — над геном коротких крыльев (v). Если муху, являющуюся доминантной гомозиготой по этим двум признакам — серую с нормальными крыльями (BBVV), скрестить с рецессивной гомозиготой по этим же признакам — черной с короткими крыльями(bbvv), то в F1 все потомство оказывается фенотипически единообразным — серое тело и длинные крылья, — а по генотипу дигетерозиготным (BbVv). На данном этапе никаких отличий от результатов обычного дигибридного скрещивания не наблюдалось (рис. 14).

Рис. 14. Сцепленное наследование генов окраски тела и длины крыльев у мухи дрозофилы
Как определить, какие гаметы образуют особи первого гибридного поколения? Для этого надо провести анализирующее скрещивание, т. е. скрещивание гибрида с рецессивной гомозиготой (в данном случае bbvv — особями, имеющими черное тело и короткие крылья), а затем посмотреть расщепление потомства от этого скрещивания (F2). Т. Морган провел две серии экспериментов по анализирующему скрещиванию. В первом случае в анализирующие скрещивания он вовлекал гибридных самцов, а во втором — гибридных самок. Рассмотрим сначала первую серию экспериментов по анализирующему скрещиванию (рис. 14, а). Гибридный самец, имевший серое тело и длинные крылья, был скрещен с самкой, имевшей черное тело и короткие крылья. Если бы гены, отвечающие за окраску тела и длину крыльев, находились в разных хромосомах, то у такого дигетерозиготного самца (BbVv) должны были бы образовываться гаметы четырех типов в равных соотношениях: 25% гамет BV, 25% bV, 25% Bv и 25% гамет bv. Следствием этого должно было бы быть появление в потомстве этого анализирующего скрещивания (в F2)четырех фенотипических классов потомков: серых длиннокрылых, серых короткокрылых, черных длиннокрылых и черных короткокрылых; при этом соотношение между классами должно было быть 1:1:1:1, т. е. по 25% особей каждого класса (см. «Анализирующее скрещивание», случай дигетерозиготы). Однако в действительности наблюдалось другое распределение: 50% серых длиннокрылых и 50% черных короткокрылых, т. е. фенотипы потомков полностью совпали с фенотипами родителей. Это значит, что в данном случае не происходило образования гамет предполагаемых четырех типов и, следовательно, гены этих признаков не находились в разных хромосомах. Иными словами, результаты этой серии экспериментов привели Т. Моргана к выводу о том, что гены цвета тела и длины крыльев находятся в одной хромосоме и поэтому следующему поколению будут передаваться сцепленно. При абсолютном сцеплении обоих генов следует ожидать не четырех (как при независимом наследовании генов), а всего лишь двух типов гамет: 50% BV и 50% bv. Это предположение подтвердилось во всех опытах, где самка была гомозиготна по обоим рецессивам (bbvv), а самец — дигетерозиготным (BbVv).
Во второй серии экспериментов родительские формы поменяли местами — дигетерозиготой (BbVv) была самка (а не самец, как раньше), а гомозиготой по обоим рецессивным аллелям(bbvv) — самец. В результате было получено совершенно иное, чем в предыдущем опыте, распределение фенотипов в потомстве.
Итак, родительские формы поменяли местами: гибридную серую длиннокрылую самку (BbVv) скрещивали с черным короткокрылым самцом (bbvv) (рис. 14, б). В потомстве получили не два, как ранее, а четыре класса фенотипов. Это были как раз те четыре фенотипа, которые следовало бы ожидать в случае независимого наследования признаков, однако между ними не наблюдалось соотношения 1:1:1:1 (или 25% :25% :25% :25%). В эксперименте было получено 41,5% серых длиннокрылых мух (BbVv), 41,5% черных короткокрылых (bbvv), 8,5% черных длиннокрылых (bbVv) и 8,5% серых короткокрылых (Bbvv). Для того чтобы у потомства сформировались данные фенотипы, самка должна образовывать не два (как в первом опыте), а четыре типа соответствующих гамет: BV, bv, bV, Bv. Откуда же взялись гаметы bV и Bv? Вспомним мейоз. В профазе первого мейотического деления происходит конъюгация гомологичных хромосом, а затем и обмен их гомологичными участками — кроссинговер. Как раз в результате этих процессов происходит перекомбинация генов, и появляются гаметы bV и Bv, т. е. гаметы с комбинациями генов, отличными от исходных групп сцепления. Такие гаметы называются кроссоверными, а гаметы, несущие исходные группы сцепления, — некроссоверными. (У самцов дрозофилы кроссоверные гаметы не образуются, потому что у них отсутствует специфический белковый комплекс, необходимый для конъюгации гомологичных хромосом.) Вторая серия опытов показала, что сцепление генов, локализованных в одной хромосоме, может быть неполным и нарушаться в результате кроссинговера.
Анализ и обобщение результатов, полученных в описанных сериях экспериментов, позволили Т. Моргану сформулировать хромосомную теорию наследственности.
Словарь основных терминов и понятий
Аморфные мутации (греч. «а» — отрицание, «морфа» — форма) — группа мутаций по характеру их проявления в фенотипе. Неактивны в отношении типичного эффекта нормального аллеля. Например, ген альбинизма полностью тормозит образование пигмента у животных или хлорофилла у растений.
Анеуплоидия (гетероплоидия) — изменение числа хромосом, не кратное гаплоидному набору. В результате возникают особи с аномальным числом хромосом: моносомики (2n - 1), у которых не хватает одной хромосомы в какой-либо паре, и полисомики, у которых одна из хромосом может быть повторена несколько раз (например, трисомики — 2n + 1, тетрасомики — 2n + 2). У человека описаны трисомии по различным парам хромосом. Так, одна добавочная хромосома у человека вызывает болезнь Дауна — физическую и умственную отсталость; недостаток одной Х-хромосомы у женщин приводит к потере признаков пола (моносомия).
Антиморфные мутации (греч. «анти» — против, «морфа» — форма) — группа мутаций по характеру их проявления в фенотипе. Оказывают действие, противоположное действию нормального аллеля. Так, у кукурузы исходный аллель дает пурпурную окраску семян, а мутантный — вызывает образование бурого пигмента.
Вариационная кривая — графическое выражение изменчивости признака, отражающее как размах вариации, так и частоту встречаемости отдельных вариант.
Вариационный ряд — ряд модификационной изменчивости признака, слагающийся из отдельных значений, расположенных в порядке увеличения или уменьшения количественного выражения признака (размеры листьев, число цветков в колосе, изменение окраски шерсти).
Гаплоидия — уменьшение числа хромосом вдвое. Такой организм (гаплоид) имеет в соматических клетках гаплоидный набор хромосом. Поскольку он имеет лишь по одной хромосоме из каждой гомологичной пары, то в его фенотипе проявляются все имеющиеся рецессивные аллели.
Генетика популяций — раздел генетики, изучающий вопросы изменения генетической структуры популяций в результате действия мутационного процесса, отбора, случайного изменения частот генов. Одна из сфер приложения популяционной генетики — теория эволюции. Фундаментальное значение для развития популяционной генетики имели два события: открытие закона Харди—Вайнберга и публикация работы С. С. Четверикова «О некоторых моментах эволюционного процесса с точки зрения современной генетики» (1926). В этой работе Четвериков показал, что рецессивная мутация, не проявляющаяся в фенотипе, тем не менее не исчезает, а накапливается в популяции в значительном количестве в гетерозиготном состоянии. Он сделал важный вывод о насыщенности природных популяций рецессивными мутациями, о том, что «популяция, подобно губке, впитывает рецессивные мутации, оставаясь при этом фенотипически однородной». Такая генетическая гетерогенность является скрытым резервом наследственной изменчивости и создает возможность эволюционных преобразований популяций под действием естественного отбора.
Генетико-автоматичёские процессы — см. Дрейф генов.
Генные (точковые) мутации — мутации, изменяющие последовательность нуклеотидов в гене, т. е. структуру самого гена.
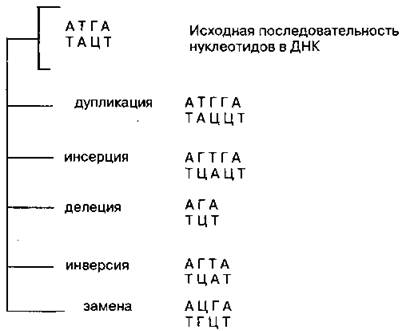
1. Генные дупликации — удвоение пары или нескольких пар нуклеотидов (удвоение пары Г—Ц).
2. Генные инсерции — вставка пары или нескольких пар нуклеотидов (вставка пары Г—Ц между А—Т и Т—А).
3. Генные делеции — выпадение нуклеотидов (выпадение комплементарной пары Т—А между А—Т и Г—Ц).
4. Генные инверсии — перестановка фрагмента гена (во фрагменте исходная последовательность нуклеотидов Т—А, Г—Ц заменяется на обратную Г—Ц, Т—А).
5. Замены нуклеотидов — замена пары нуклеотидов на другую; при этом общее число нуклеотидов не меняется (замена Т—А на Ц—Г). Один из наиболее частых типов мутаций.
Дупликации, инсерции и делеции могут приводить к изменению рамки считывания генетического кода. Рассмотрим это на примере. Возьмем следующую исходную последовательность нуклеотидов в ДНК (для простоты будем рассматривать только одну ее цепь): АТГАЦЦГЦГА... Она будет считываться следующими триплетами: АТГ, АЦЦ, ГЦГ, А... Допустим, произошла делеция, и в самом начале последовательности между А и Г выпал нуклеотид Т. В результате этой мутации получится измененная последовательность нуклеотидов: АГАЦЦГЦГА, которая уже будет считываться совершенно иными триплетами: АГА, ЦЦГ, ЦГА. Поэтому в полипептидную цепь будут соединяться совершенно другие аминокислоты и, таким образом, будет синтезироваться мутантный белок, совершенно непохожий на нормальный. Кроме того, в результате генных мутаций, приводящих к сдвигу рамки, могут образовываться терминирующие кодоны ТАА, ТАГ или ТГА, прекращающие синтез. Выпадение целого триплета приводит к менее тяжелым генетическим последствиям, чем выпадение одного или двух нуклеотидов. Рассмотрим ту же нуклеотидную последовательность: АТГАЦЦГЦГА... Допустим, произошла делеция, и выпал целый триплет АЦЦ. Мутантный ген будет иметь измененную последовательность нуклеотидов АТГГЦГА, которая будет считываться следующими триплетами: АТГ, ГЦГ, А... Видно, что после выпадения триплета рамка считывания не сдвинулась, синтезированный белок хоть и будет на одну аминокислоту отличаться от нормального, но в целом будет весьма на него похож. Однако это отличие в аминокислотном составе может привести к изменению третичной структуры белка, которая в основном и определяет его функции, и функция мутантного белка, скорее всего, будет снижена по сравнению с нормальным белком. Этим и объясняется тот факт, что мутации, как правило, рецессивны.
Генные мутации проявляются фенотипически в результате синтеза соответствующих белков:
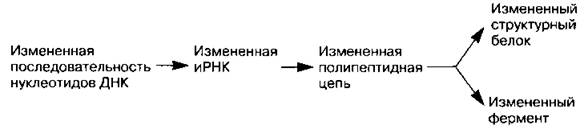
Генные мутации приводят к изменению строения молекул белков и к появлению новых признаков и свойств (например, альбиносы у животных и растений, махровость у цветков за счет преобразования тычинок в лепестки и снижение их плодовитости, образование летальных и полулетальных генов, вызывающих гибель организма, и т. д.). Генные мутации происходят под влиянием мутагенных факторов (биологических, физических, химических) или спонтанно (случайно). Генные мутации свойственны и генетической РНК вирусов.
Геномные мутации — изменение числа хромосом. Они могут быть вызваны нерасхождением хромосом при мейозе, что приводит к появлению у гамет нового набора хромосом. Геномные мутанты могут быть представлены гаплоидами (с вдвое меньшим числом хромосом), анеуплоидами (с лишней или недостающей хромосомой), полиплоидами (с кратным увеличением наборов хромосом).
Гипоморфные мутации — группа мутаций по характеру их проявления. Действуют в том же направлении, что и нормальный аллель, но дают несколько ослабленный эффект. Например, у дрозофилы окраска глаз при мутации значительно бледнее.
Гомологические ряды (греч. «гомологос» — подобный) — сходные, параллельные ряды наследственных изменений, характерные для родственных видов и родов. Закон гомологических рядов наследственной изменчивости был открыт Н. И. Вавиловым в 1920 г. — «Изменчивость близких по происхождению видов и родов осуществляется общим путем. Генетически близкие роды и виды характеризуются сходными рядами наследственной изменчивости с такой правильностью, что, зная ряд форм в пределах одного вида, можно предвидеть нахождение параллельных форм у других родственных видов и родов. Чем более систематически близки виды и роды, тем полнее сходство в рядах их изменчивости». Согласно закону наличие ряда признаков у одного вида предполагает наличие таких же признаков и у другого родственного вида. Так, отсутствие ядовитых растений у подавляющего большинства представителей семейства бобовых привело ученого к мысли о том, что у ядовитого люпина имеются в природе неядовитые формы. В результате поисков такой люпин был найден. Таким же образом была найдена одноростковая свекла, безостая твердая пшеница. Закон имеет большое практическое значение, так как дает возможность селекционеру искать или создавать формы, известные у других видов или родов. Аналогичное явление характерно и для животных. Например, селекционным путем была получена вся гамма расцветок меха у норки (34 типа).
Дизруптивный (рассекающий) отбор — одна из форм естественного отбора, которую можно рассматривать как противоположность стабилизирующему. Дизруптивный отбор действует тогда, когда условия внешней среды настолько изменились, что основное число особей вида утрачивает адаптивность, а преимущества приобретают особи с крайними отклонениями от средней нормы. Именно эти отклонения и играют адаптивную роль в новых условиях. Численность таких форм быстро увеличивается, и на базе одного вида вследствие дивергенции формируются несколько новых.
Дрейф генов — случайное ненаправленное изменение частот аллелей и генотипов в малых популяциях. Одной из причин дрейфа генов могут быть популяционные волны. Поскольку дрейф генов может идти вопреки естественному отбору, его последствия для популяции непредсказуемы: небольшую популяцию он может привести к гибели либо может сделать ее еще более приспособленной к данной среде или усилить ее отличие от родительской популяции. Дрейф генов считают существенным фактором в возникновении новых видов в островных изолированных популяциях. Явление дрейфа генов было открыто С. Райтом и Р. Фишером и независимо от них советскими учеными Н. П. Дубининым и Д. Д. Ромашовым, которые дали ему название «генетико-автоматические процессы».
Закон Харди—Вайнберга — основа математических построений генетики популяций и современной эволюционной теории. Сформулирован независимо друг от друга математиком Г. Харди (Англия) и врачом В. Вайнбергом (Германия) в 1908 г. Этот закон утверждает, что частоты аллелей и генотипов в данной популяции будут оставаться постоянными из поколения в поколение при выполнении следующих условий: 1) численность особей популяции достаточно велика (в идеале — бесконечно велика), 2) спаривание происходит случайным образом (т. е. осуществляется панмиксия), 3) мутационный процесс отсутствует, 4) отсутствует обмен генами с другими популяциями, 5) естественный отбор отсутствует, т. е. особи с разными генотипами одинаково плодовиты и жизнеспособны. Иногда этот закон формулируют иначе: в идеальной популяции частоты аллелей и генотипов постоянны. (Поскольку описанные выше условия выполнения данного закона и есть свойства идеальной популяции.) Математическая модель закона отвечает формуле:
р2 + 2pq + q2 = 1
Она выводится на основе следующих рассуждений. В качестве примера возьмем простейший случай — распределение двух аллелей одного гена. Пусть два организма являются основателями новой популяции. Один из них является доминантной гомозиготой (АА), а другой — рецессивной гомозиготой (аа). Естественно, что все их потомство в F1 будет единообразным и будет иметь генотип (Аа). Далее особи Fj будут скрещиваться между собой. Обозначим частоту встречаемости доминантного аллеля (А) буквой р, а рецессивного аллеля (а) — буквой q.Поскольку ген представлен всего двумя аллелями, то сумма их частот равна единице, т. е. р -I- q = 1. Рассмотрим все яйцеклетки в данной популяции. Доля яйцеклеток, несущих доминантный аллель (А), будет соответствовать частоте этого аллеля в популяции и, следовательно, будет составлять р. Доля яйцеклеток, несущих рецессивный аллель (а), будет соответствовать его частоте и составлять q. Проведя аналогичные рассуждения для всех сперматозоидов популяции, придем к заключению о том, что доля сперматозоидов, несущих аллель (А), будет составлять р, а несущих рецессивный аллель (а) — q. Теперь составим решетку Пеннета; при этом при написании типов гамет будем учитывать не только геномы этих гамет, но и частоты несомых ими аллелей. На пересечении строк и столбцов решетки мы получим генотипы потомков с коэффициентами, соответствующими частотам встречаемости этих генотипов.
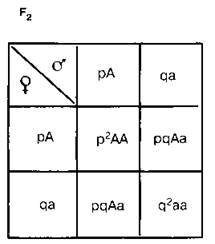
Из приведенной решетки видно, что в F2 частота доминантных гомозигот (АА) составляет р2, частота гетерозигот (Аа) — 2pq, а рецессивных гомозигот (аа) — q2. Поскольку приведенные генотипы представляют собой все возможные варианты генотипов для рассматриваемого нами случая, то сумма их частот должна равняться единице, т. е.
р2 + 2pq + q2 = 1
Главное применение закона Харди—Вайнберга в генетике природных популяций — вычисление частот аллелей и генотипов. Рассмотрим пример использования этого закона в генетических расчетах. Известно, что один человек из 10 тыс. является альбиносом, при этом признак альбинизма у человека определяется одним рецессивным геном. Давайте вычислим, какова доля скрытых носителей этого признака в человеческой популяции. Если один человек из 10 тыс. является альбиносом, то это значит, что частота рецессивных гомозигот составляет 0,0001, т. е. q2= 0,0001. Зная это, можно определить частоту аллеля альбинизма q, частоту доминантного аллеля нормальной пигментации р и частоту гетерозиготного генотипа (2pq). Люди с таким генотипом как раз и будут скрытыми носителями альбинизма, несмотря на то что фенотипически этот ген не будет у них проявляться и они будут иметь нормальную пигментацию кожи.
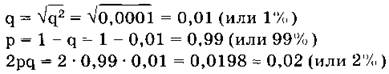
Из приведенных простых расчетов видно, что, хотя число альбиносов крайне невелико — всего лишь один человек на 10 тыс., ген альбинизма несет значительное количество людей — около 2%. Иными словами, даже если признак фенотипически проявляется очень редко, то в популяции присутствует значительное количество носителей этого признака, т. е. особей, имеющих этот ген в гетерозиготе.
Благодаря открытию закона Харди—Вайнберга процесс микроэволюции стал доступен непосредственному изучению: о его ходе можно судить по изменениям из поколения в поколение частот генов (или генотипов). Таким образом, несмотря на то что этот закон действителен для идеальной популяции, которой нет и не может быть в природе, он имеет огромное практическое значение, так как дает возможность рассчитать частоты генов, изменяющиеся под влиянием различных факторов микроэволюции.
Запас изменчивости в популяции — накопившийся в многочисленных поколениях резерв признаков наследственной изменчивости, находящихся в гетерозиготном состоянии. С того момента, когда они переходят в гомозиготное состояние и проявляются фенотипически, они попадают под контроль естественного отбора. В природных популяциях мушки дрозофилы выявлено, что каждая особь гетерозиготна по одной морфологической, двум летальным и четырем полулетальным мутациям. Этот генетический груз составляет тот резерв наследственной изменчивости данного вида, который проявляется при изменении среды обитания и поставляет материал для естественного отбора. На одного человека в среднем приходится до 10 рецессивных летальных мутаций. В связи с мутагенным действием загрязнения окружающей среды груз летальных мутаций возрастает, поэтому участились случаи рождения детей с тяжелыми наследственными пороками (как умственными, так и физическими).
Изменчивость — способность организмов изменять свои признаки и свойства, что проявляется в разнообразии особей внутри вида. Изменчивость бывает наследственная (неопределенная, индивидуальная генотипическая) и ненаследственная (определенная, групповая, модификационная). Наследственная изменчивость связана с изменением генотипа, ненаследственная — с изменением фенотипа под влиянием условий окружающей среды.
Изменчивость при обмене генетической информацией — наследственные изменения, возникающие в результате встраивания в генетический аппарат про- или эукариотической клетки фрагмента чужеродной ДНК, несущего определенные гены. В отличие от мутагенеза (когда в результате какого-либо воздействия может измениться любой ген или несколько генов одновременно и результат этих изменений невозможно предсказать заранее) при обмене генетической информацией происходит направленное изменение определенных генов, и результат этих изменений заранее известен. Обмен генетической информацией осуществляется в виде трансформации и трансдукции. Используется в генной инженерии.
Комбинационная (комбинативная) изменчивость — тип наследственной изменчивости, обусловленной различными перекомбинациями уже имеющихся генов и хромосом. Не сопровождается изменениями структуры генов и хромосом. Ее источниками являются следующие процессы, происходящие в ходе мейоза и в результате оплодотворения:
— рекомбинация генов в результате кроссинговера;
— рекомбинация хромосом в ходе мейоза;
— комбинации хромосом, возникающие в результате слияния гамет при оплодотворении.
Модификации — изменения фенотипа (признаков организма), вызванные факторами внешней среды и не связанные с изменениями генотипа организма. Модификации являются изменениями в пределах нормы реакции, которая контролируется генотипом. Модификации характеризуются рядом свойств:
— возникают постепенно, имеют переходные формы;
— модификации — изменения количественные; они образуют непрерывные ряды и группируются вокруг среднего значения;
— возникают направленно — под влиянием одного и того же фактора среды группа организмов изменяется сходным образом;
— модификации обратимы;
— модификации не передаются по наследству из поколения в поколение.
Основой существования модификаций является то, что фенотип — это результат взаимодействия генотипа и внешних условий, поэтому изменение внешних условий может вызвать изменения фенотипа, не затрагивая генотип. Эволюционное значение модификаций заключается в том, что они позволяют организмам адаптироваться к условиям внешней среды, которые могут неоднократно меняться в течение жизни этих организмов.
Модификационная изменчивость — изменчивость фенотипа, которая является реакцией конкретного генотипа на изменившиеся условия среды. Модификационные изменения не передаются по наследству и возникают как реакция организма, т. е. представляют собой адаптацию. Так, при ежедневном обливании холодной водой появляется устойчивость к простудным заболеваниям, так как вследствие закаливания организма клеточные мембраны не теряют свойств избирательной проницаемости при низких температурах и оказывают сопротивление микроорганизмам, которые всегда находятся в окружающей среде. Если человек прекращает закаливание, организм вновь подвергается простудным заболеваниям. В практике сельского хозяйства модификационная изменчивость проявляется в изменении продуктивности животных и растений. Так, дополнительное освещение на птицефермах приводит к повышению яйценоскости кур, поскольку создаются условия длинного естественного дня, при котором куры несутся чаще. Биологические стимуляторы способствуют увеличению удойности коров и повышению жирности молока. Своевременные агротехнические мероприятия (полив, культивация, внесение удобрений) значительно повышают урожайность сельскохозяйственных культур. Модификационная изменчивость ограничена пределами, которые допускает норма реакции генотипа особи.
Мутагены (лат. «мутацио» — изменение и греч. «генос» — рождение) — факторы, вызывающие изменение генотипа как в естественных, так и в искусственно созданных условиях. Это могут быть химические, температурные, биологические воздействия, действия различных излучений и др. Например, загрязнение атмосферы и воды отходами химической промышленности, которые являются мутагенами, приводит к заболеванию или гибели живых организмов. Пестициды, вызывая мутации и гибель организмов, против которых они направлены, одновременно оказывают вредное воздействие на другие организмы, в том числе и на человека. Биологическими мутагенами являются вирусы и бактерии (ДНК вирусов встраивается в клетку и разрушает или изменяет ее генотип). Химическими мутагенами служат некоторые лекарства, возбуждающие средства (алкоголь), наркотики (никотин табака, морфин мака). Особенно губительны эти мутагены для детей и молодых людей.
Мутации — внезапные, естественные или вызванные искусственно наследуемые изменения генетического материала, приводящие к изменению признаков организма. Основы учения о мутациях заложены Г. де Фризом в 1901 г. и оформлены затем в мутационную теорию. Мутации характеризуются рядом свойств:
— возникают внезапно, скачкообразно, без всяких переходных форм;
— мутации — изменения качественные и, в отличие от ненаследственных изменений, не образуют непрерывных рядов и не группируются вокруг среднего значения;
— возникают ненаправленно — под влиянием одного и того же мутагенного фактора может мутировать любая часть структуры, несущей генетическую информацию, приводя тем самым к изменению самых разнообразных признаков;
— сходные мутации могут возникать неоднократно;
— мутации передаются из поколения в поколение.
Мутационная изменчивость — тип наследственной изменчивости, обусловленной появлением различных изменений в структуре генов, хромосом или генома. Фенотипически мутации проявляются только тогда, когда становятся гомозиготными.
Направленное получение мутаций — новый подход в генетике и молекулярной биологии, базирующийся на раскрытии молекулярных механизмов мутаций, управлении этими мутациями и получении форм живых организмов с заданными признаками и свойствами. В селекции растений — это выведение полиплоидных форм, в микробиологии — новых штаммов микроорганизмов, способных синтезировать определенные органические вещества (кормовые или лекарственные).
Неоморфные мутации (греч. «неос» — новый, «морфа» — форма) — группа мутаций, нетипичных по характеру их проявления в фенотипе. Их действие совершенно отлично от действия исходного нормального аллеля.
Норма реакции — предел модификационной изменчивости признака, обусловленный генотипом. Качественные признаки, такие, как окраска шерсти животных, цветка растений, имеют узкий предел изменчивости, поскольку являются жизненно важными (окраска животных играет защитную роль, окрашенный венчик цветка привлекает насекомых-опылителей). Количественные признаки, например масса животного, размеры листьев растения, изменяются в довольно широких пределах. Однако такие количественные признаки, как размеры сердца и мозга, имеют узкую норму реакции. Норма реакции выражается в виде вариационного ряда или в виде вариационной кривой.
Полиплоидия (греч. «поли» — много, «плоид» — складывать) — увеличение числа хромосом, кратное (эуплоидия) и некратное (анэуплоидия) гаплоидному набору: 3n — триплоид, 4n — тетраплоид, 5n — пентаплоид и т. д. Причина возникновения — удвоение хромосом без последующего деления клетки либо нерасхождение хромосом в результате блокады работы веретена деления. Среди животных встречается крайне редко (тутовый шелкопряд), так как, как правило, приводит к гибели организма. У растений распространено очень широко. В пределах одного рода различные виды часто образуют полиплоидные ряды. Так, род картофель имеет виды с 12, 24, 36, 48, 60, 72, 96, 108 и 144 хромосомами. Полиплоидные растения имеют более широкую норму реакции и, следовательно, легче приспосабливаются к неблагоприятным условиям внешней среды. В северных широтах и высокогорных районах полиплоиды составляют более 80% от общего числа распространенных там видов растений. Полиплоидные формы известны в декоративном цветоводстве, например тюльпаны, нарциссы, гладиолусы, имеющие очень крупные цветки. Некоторые полиплоиды имеют пониженную семенную продуктивность вследствие того, что у их триплоидных форм при мейозе часть хромосом не находит себе пары. При этом образуются половые клетки с несбалансированным набором хромосом, которые затем погибают, что приводит к снижению плодовитости или к полной бесплодности (так могут быть получены бессемянные плоды). В тетраплоидном же организме набор хромосом обычно бывает парным и мейоз идет нормально, что обеспечивает высокую семенную продуктивность растений.
Полиплоидные ряды — формы растений с удвоенным, утроенным и т. д. по сравнению с исходным видом набором хромосом. В селекционной практике получены полиплоидные ряды свеклы, пшеницы, кукурузы, гречихи, картофеля, плодовых культур, дающие большую прибавку урожая и отличающиеся большой жизнеспособностью.
Популяционные волны (волны численности, волны жизни) — резкие колебания численности особей популяции вследствие естественных причин. Впервые на это явление обратил внимание С. С. Четвериков, им же был введен данный термин (1905). У быстро размножающихся видов наблюдается периодическое чередование быстрых подъемов численности и резких спадов. Например, численность мышевидных грызунов за четыре, года возрастает от ничтожной до максимальной, затем падает почти до нуля и снова начинает увеличиваться. Важную роль в данном случае играют хищники, численность которых колеблется пропорционально численности грызунов. Важной причиной популяционных волн является также постепенное истощение пищевых ресурсов в связи с ростом популяции и последующее их восстановление после снижения численности популяции. Причинами резких непериодических снижений численности популяции могут также быть стихийные бедствия: засухи, пожары, наводнения. Каким бы ни был механизм популяционных волн, ясно, что на численность популяции могут влиять одновременно многие факторы. Популяционные волны играют большую роль в ходе микроэволюции. С возрастанием численности популяции увеличивается вероятность появления новых мутаций и их комбинаций. Если в среднем один мутант появляется на 10 тыс. особей, то при возрастании численности популяции в 100 раз общее число мутантов увеличится во столько же раз. После спада волны численности генофонд популяции может уже оказаться иным: часть мутаций может случайно исчезнуть из-за гибели несущих их особей, а частота встречаемости других мутаций может повыситься. Таким образом, популяционные волны сами по себе не вызывают наследственную изменчивость, а только способствуют изменению частот аллелей и генотипов; они являются поставщиком исходного материала для действия естественного отбора.
Репарация — исправление мутационных повреждений генов; процесс, противоположный мутагенезу. В ДНК клетки ежедневно происходит множество случайных изменений. Однако из тысяч таких случайных замен оснований в ДНК лишь одна приводит к мутации, а все остальные эффективно исправляются (репарируются) с помощью специальных ферментных систем. Такие ферментные системы выработались в ходе эволюции и направлены на поддержание стабильности генетической информации клетки. Репарация может происходить как до, так и после репликации ДНК. Конкретные механизмы репарации различны, но все они основаны на том, что в молекуле ДНК имеется две копии информации (в двух комплементарных цепях нуклеотидов). Репарация обязательно включает в себя следующие три этапа:
— распознавание и удаление поврежденного участка, образование бреши в одной из цепей ДНК;
— заполнение этой бреши: синтез на этом месте нового фрагмента полинуклеотидной цепи. Синтез идет по принципу комплементарности, в качестве матрицы используется вторая неповрежденная цепь молекулы ДНК;
— «пришивание» вновь синтезированного фрагмента к полинуклеотидной цепи и восстановление структуры молекулы.
Наиболее хорошо механизмы репарации изучены на микроорганизмах.
Синтетическая теория эволюции — это современная эволюционная теория, основы которой были заложены С. С. Четвериковым (1926), объединившим дарвинизм с классической генетикой. Эта теория получила развитие в работах английского ученого Дж. Хаксли (1942), который продолжил разработку комплексного подхода к процессам эволюции на базе современных достижений генетики популяций, молекулярной биологии, эволюции биосферы. Основными положениями этой теории являются следующие:
1. Популяция — наименьшая, элементарная эволюционная единица.
2. Элементарным эволюционным событием является изменение генетического состава популяции.
3. Материалом для эволюции служат, как правило, мелкие, дискретные изменения наследственности — мутации.
4. Факторами, поставляющими материал для отбора, являются мутационный процесс, комбинативная изменчивость и волны численности (популяционные волны). Они имеют случайный и ненаправленный характер. Фактором, усиливающим генетические различия, является изоляция.
5. Единственный направляющий фактор эволюции — естественный отбор, возникающий на основе борьбы за существование. Его действие основывается на сохранении и накоплении случайных мелких мутаций.
6. Эволюция — постепенный и длительный процесс. Видообразование как этап эволюционного процесса представляет собой последовательную смену временных популяций.
7. Вид, как правило, состоит из множества соподчиненных, морфологически, физиологически и генетически отличных, но репродуктивно не изолированных единиц — подвидов и популяций. (Однако известны также виды с небольшими ареалами, не делящиеся на подвиды, а некоторые реликтовые виды состоят из единственной популяции.)
8. Обмен аллелями, поток генов возможны лишь внутри вида. (Отсюда следует определение вида как генетически целостной и замкнутой системы.)
9. Эволюция носит дивергентный характер, т. е. один таксон может стать предком нескольких дочерних таксонов, но каждый вид имеет единственный предковый вид, единственную предковую популяцию. (Вследствие этого любой реальный, а не сборный таксон имеет монофилетическое происхождение.)
10. Макроэволюция на уровне выше вида (род, семейство, отряд и т. д.) идет лишь путем микроэволюции; не существует закономерностей макроэволюции, отличных от закономерностей микроэволюции.
11. Эволюция непредсказуема и имеет ненаправленный характер, т. е. не идет в направлении какой-то конечной, заранее заданной цели. Иными словами, она не носит финалистического характера.
«Классическая» синтетическая теория эволюции сформировалась в 40-х гг. XX в. Современная эволюционная биология дополняет ее рядом положений:
— Помимо естественного отбора, в небольших изолированных популяциях формирующую роль играет дрейф генов.
— Эволюция не всегда совершается постепенно. Видообразование может носить и внезапный характер, например путем полиплоидии или в результате крупных хромосомных мутаций.
— Эволюция может быть прогнозируема: в ряде случаев ее общее направление можно предсказать, оценивая прошлую историю вида, его генофонд и возможное влияние среды.
Спонтанные мутации (лат. «спонтанус» — самопроизвольный) — мутации, возникающие в естественных условиях. Они носят случайный, ненаправленный характер и могут происходить как в половых, так и в соматических клетках (клетках тела). Причины спонтанных мутаций — нерасхождение хромосом при делении клеток, перестройка хромосом, изменение последовательности нуклеотидов в ДНК, перестройка генов и т. д. Спонтанные мутации — важнейший источник наследственных изменений, материал для естественного отбора, элементарное эволюционное явление.
Точковые мутации — см. Генные (точковые) мутации.
Трансдукция — направленный перенос и встраивание в генетический аппарат клетки небольшого фрагмента чужеродного ДНК при помощи посредников — бактериофагов или вирусов. Открыта в 1952 г. Н. Циндером и Дж. Ледербергом. В одной из клеток в геном вируса встраивается фрагмент генетического материала бактерии, после чего вирусные частицы проникают в другую клетку и встраивают в ее геном уже не только свою нуклеиновую кислоту, но и приобретенный фрагмент ДНК. После выделения вирусной нуклеиновой кислоты в геноме этой другой клетки остается фрагмент генетического материала первой бактерии. Трансдукция используется в генной инженерии.
Трансформация — направленный перенос и встраивание в генетический аппарат клетки небольшого фрагмента чужеродной ДНК. Происходит без участия вирусов и бактериофагов. В ходе этого процесса небольшой фрагмент ДНК поглощается клеткой- реципиентом и включается в состав своей собственной ДНК, замещая в ней похожий (но не обязательно идентичный) фрагмент. Наблюдается лишь у немногих бактерий. Трансформация была открыта в 1944 г. О. Эвери, С. Мак-Леодом и М. Мак-Карти. Открытие этого явления было первым прямым доказательством того, что генетическим материалом является именно ДНК (а не белки, как предполагалось ранее).
Хромосомные аберрации — см. Хромосомные мутации.
Хромосомные мутации (аберрации, перестройки) — изменения положения участков хромосом; приводят к изменению размеров и формы хромосом. В эти изменения могут быть вовлечены как участки одной хромосомы, так и участки разных, негомологичных хромосом, поэтому хромосомные мутации (перестройки) подразделяются на внутри- и межхромосомные.
А. Внутрихромосомные мутации
1. Хромосомные дупликации — удвоение участка хромосомы.
2. Хромосомные делеции — утрата хромосомой какого-либо участка.
3. Хромосомные инверсии — разрыв хромосомы, переворачивание оторвавшегося участка на 180° и встраивание его на прежнее место.

Б. Межхромосомные мутации
1. Транслокация — обмен участками между негомологичными хромосомами (в мейозе).
2. Транспозиция — включение участка хромосомы в другую, негомологичную хромосому без взаимного обмена.
Хромосомные перестройки — см. Хромосомные мутации.
Цитоплазматическая мужская стерильность (ЦМС) — разновидность внеядерной наследственности. Характеризуется присутствием в ДНК митохондрий и пластид цитоплазматического гена (плазмогена), угнетающего формирование хромосом при образовании пыльцы. В результате появляется нежизнеспособная (стерильная) пыльца, не образующая спермиев. ЦМС играет большую роль в селекции и семеноводстве для исключения самоопыления и последующего получения гетерозисных гибридов. Она описана у кукурузы, лука, пшеницы, свеклы и других растений.
Цитоплазматическая (внеядерная) наследственность — наследственность, осуществляемая с помощью молекул ДНК, находящихся в пластидах и митохондриях. В этом случае признаки не наследуются по законам Менделя, так как указанные молекулы ДНК находятся вне хромосом. Признаки, наследующиеся цитоплазматически, передаются только через яйцеклетку, содержащую зачатки пластид и митохондрий, например пестролистность растения львиный зев передается по материнской линии, так как яйцеклетка содержит хлоропласты в виде пропластид, спермий же пластид не содержит. Пластиды размножаются делением, но поскольку не все они содержат хлорофилл (в силу мутаций), растение получается пестролистным. У человека также встречается цитоплазматическое наследование признаков, например одна из форм несращения остистых отростков позвонков.