Биогеография общая и частная: суши, моря и континентальных водоёмов - 2017 год
Крупные озёра - Причины и скорость образования новых видов и экосистем - Био-экология, или синэкология
Многим крупным и даже не очень крупным озёрам свойственен эндемизм. Байкал — классический в отечественной биогеографической литературе пример региона с высоким эндемизмом. История Байкала рассмотрена в Главе 4. Широко распространено мнение, связывающее высокий эндемизм байкальской биоты её древностью. Одни объясняют отличия байкальской фауны, её обособленность происхождением от фауны мезозойских морских бассейнов Азии. Другие находят, что обособленность фауны Байкала от окружающей связана с вымиранием последней, т. е. считают байкальскую фауну реликтовой. Общим для этих двух точек зрения является то, что их сторонники убеждены, что фауна Байкала богата эндемиками потому, что она остается реликтом какой-то ещё более богатой фауны, а сам Байкал рассматривают как рефугиум. Поэтому их можно объединить в одну гипотезу реликтов.
“Предполагается, что причиной несмешиваемости фаун Байкала и окружающих водоёмов служит особое строение байкальской воды, содержащей примесь изотопических молекул, увеличивающих её плотность” (Бирштейн, 1946: 244).
“Байкальская флора и фауна не смешивается с общесибирской не по причине своей древности, а по причине своей специфичности, выработанной в процессе видообразования в самом Байкале” (Абросов, 1987: 10).
Данная гипотеза в действительности объясняет отнюдь не все факты; остаётся неясным:
— почему существующие отличия Байкала и окружающих водоёмов столь резки, т. е. почему реликты сохранились почти исключительно в Байкале (а также водоёмах с условиями, сходными с байкальскими, в которые они могли попасть из Байкала)? Для объяснения этого выдвигали даже такие фантастические теории, как специфический изотопный состав байкальской воды (Верещагин, 1940б; Бирштейн, 1946; Берг, 1955).
— почему высокий эндемизм обеспечен на основе очень небольшого числа предковых форм (Табл. 1.4)?
Табл. 1.4. Число предполагаемых исходных видов и число развившихся из них таксонов в фауне Байкала. По Кожову (1962), Семерному (2004), Камалтинову (2002); Strong et al. (2008).
|
таксон |
число исходных видов |
эндемичных |
||
|
видов |
родов |
семейств (подсемейств) |
||
|
губки |
1-2 |
6 |
3 |
1 |
|
турбеллярии |
3-4 |
до 40 |
? |
- |
|
олигохеты |
? |
164 |
11 |
- |
|
остракоды |
1-2 |
19 |
- |
- |
|
изоподы (Asellus) |
2 |
5 |
||
|
Gammaridae |
4-5 |
363 |
72 |
7 |
|
ручейники |
2-3 |
10-12 |
2 |
|
|
гастроподы |
5-6 |
114 |
6 |
3 |
|
рыбы |
2-3 |
23 |
7 |
3 |
Если рассматривать проблему шире, не замыкаясь на Байкале, при анализе сходных биот, оказывается, что обе данные особенности свойственны и другим крупным озёрам с высоким эндемизмом, но совершенно другой историей. В отношении этих озёр также выдвигали теории о происхождении их биоты из морской (примеры см.: Старобогатов, 1970). Объяснить озёрный эндемизм морским происхождением пытались даже для Иссык-Куля, расположенного на высоте 1608 м над уровнем моря! Одни из таких озёр с высоким эндемизмом, как и Байкал, представляют собой трещины в коре, а другие имеют большую площадь, но сравнительно мелководны (табл. 1.5, Рис. 1.29). В качестве примера можно привести озеро Ньяса (Малави). Это озеро возникло примерно 500 тыс. лет назад (современный глубоководный режим установился там гораздо позднее — примерно 50 тыс. лет назад). Из примерно 250 видов обитающих тут рыб более 90% — эндемы: около 100 эндемичных видов широко распространённого рода цихлид Haplochromis, 70 других видов цихлид, принадлежащих к 20 эндемичным родам (Banarescu, 1995) (Рис. 1. 30) и несколько эндемичных видов рыб других семейств. По другим данным в нём обитает более 200 видов цихлид, из которых — эндемичны 180 (Махлин, 1983) или 178 видов, из которых 174 — эндемы (Дарлингтон, 1966). Эндемичные цихлиды образовались из двух исходных видов (Roberts, 1975). Впадина озера Танганьика в три раза старше (1,5—2 млн. лет), но в последние 50—60 тыс. лет уровень озера неоднократно снижался на сотни метров (до 600 м). Специфичность ихтиофауны озера примерно та же на видовом уровне и выше на родовом: из общего числа примерно 200 видов (примерно 40 родов) 95% — эндемы. Несомненно, что условия и имеющие ресурсы в этих огромных озёрах существенно отличны от таковых в окружающих мелких водоёмах и реках. Поэтому в данных озёрах не могли функционировать экосистемы окружающих водоёмов. В полном соответствии с теорией структурализма при образовании значительной территории, не могущей быть населённой ни одной из существующих экосистем, в них сформировались самостоятельные экосистемы, о чём и свидетельствует образование эндемичных для них видов. Принять участие в формировании новых биот в силу разных причин смогли немногие виды, именно поэтому многочисленные эндемичные виды образовались из немногих исходных, как, например, в Байкале (табл. 1.4). Новые виды образовывались для потребления специфических для данного озера ресурсов, в различных озёрах эти ресурсы неодинаковые, различны и донорские биоты, поэтому в каждом озере в одних группах наблюдается высокое разнообразие, тогда как другие или малоразнообразны, или вовсе отсутствуют. Группы, дающие вспышки видообразования в разных озёрах системах, различны. Так, в частности для тропических африканских озёр характерен высокий эндемизм рыб и наяд (группа двустворчатых моллюсков, куда входят беззубки и перловицы), тогда как фауна бокоплавов и изопод (таксонов разнообразие которых в тропиках гораздо ниже, чем в умеренных широтах) крайне бедна; в Байкале же, расположенном в умеренных широтах, исключительно велико разнообразие бокоплавов и изопод, эндемичных рыб немного, а эндемичных наяд в нём вовсе нет.
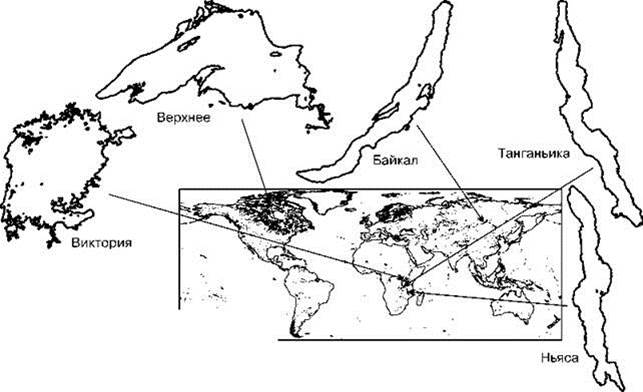
Рис. 1.29. Некоторые крупные озёра в одном масштабе.
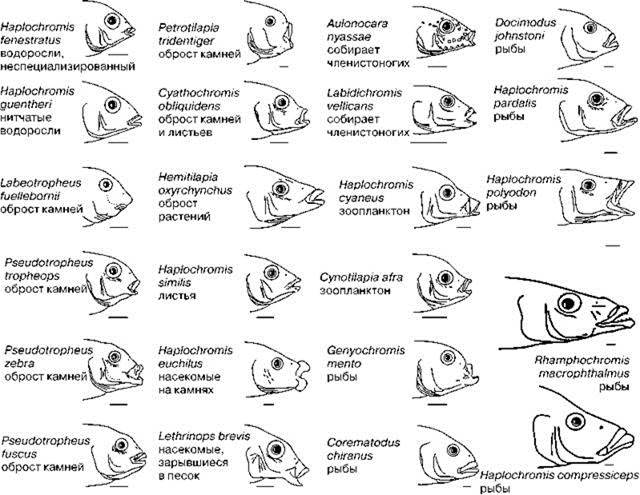
Рис. 1.30. Цихлиды оз. Ньяса. Все шкалы — 1 см. По Fryer, Iles (1972). Указан объект питания.
Табл. 1.5. Морфометрические данные некоторых крупных озёр.
|
озеро |
площадь тыс. км2 |
глубина, м |
|
|
максимальная |
аэробной зоны |
||
|
Верхнее |
82,4 |
393 |
393 |
|
Виктория |
68 |
80 |
80 |
|
Танганьика |
34 |
1470 |
70-240 |
|
Байкал |
31,5 |
1620 |
1620 |
|
Ньяса |
30,8 |
706 |
35-50 |
Скорость образования видов (эндемов) при формировании новой экосистемы поистине сказочно велика, что находится в резком противоречии с общепринятыми представлениями о скоростях видообразования и вообще об эволюции как плавном и неторопливом процессе. Так, согласно современным данным, оз. Виктория полностью высохло в позднем плейстоцене, примерно 12 тысяч лет назад (Johnson et al., 1996). Очевидно, что как минимум большая часть видов биоты существовавшего до того озера вымерла. Тем не менее, современная биота озера насчитывает много эндемов. Так, фауна цихлид составляет по разным оценкам: 160—170 видов цихлид, из них лишь три — неэндемичные (Махлин, 1983); около 200 видов, из которых 169 — эндемы (Banarescu, 1995); более 300 эндемичных видов (Brown, Lomolino, 1998). По-видимому, для образования большей части эндемичных видов оказалось достаточно этих 12 тысяч лет. Вероятно также, что видообразование происходило симпатрически без какой-либо изоляции.
Озеро Ланао (Lanao) на Филиппинах почти в 200 раз меньше озера Виктория (375 км2) и имеет примерно тот же возраст (10 тыс. лет). Herre в 1924 и 1932 гг. описал 13 эндемичных видов (но не цихлид, практически отсутствующих в Азии, а разнообразных здесь карповых Cyprinidae), относящихся к одному широко распространённому и четырём эндемичным родам, три из которых монотипичны, а один включает два вида. Данные виды несомненно произошли от одного современного вида Puntius binotatus (=Barbus binotatus), широко рапспространённого на Филиппинах. При этом у Mandibularca и Spratellicypris строение нижней челюсти выходит за пределы вариабельности во всем семействе карповых (275 родов и более 1700 видов). Их валидность одними была признана (Myers, 1960), другими — нет (Banarescu, 1992). Основной аргумент P. Banarescu, в принципе отрицающего возможность симпатрического видообразования, звучит так: “evidently not consistent with princeples of the strictly phyletical (hennigian) classification” (в вольном переводе: “этого не может быть потому что этого не может быть никогда”). Также он указывает, что три из пяти видов эндемичных родов были описаны по одному экземпляру, а голотипы утрачены во время войны. В дальнейшем, несмотря на интенсивные попытки, не удалось их поймать, правда, озеро с тех пор подверглось сильному антропогенному воздействию, и они могли вымереть (Leveque et al., 2008). В то же время P. Banarescu признаёт, что краб Archipelothelphusa wolterecki действительно эндемичен для Ланао. Относительно малакофауны озера Я.И. Старобогатов отмечает, что “почти все виды, отмеченные здесь, эндемичны для озера” (1970: 210).
У обитающих в Танганьике цихлид строение зубов также выходит за пределы вариабельности, но даже не семейства, а всего класса: у видов эндемичного Perissodus зубы двувершинные или листовидные (Рис. 1.31), приспособленные для скусывания чешуи рыб — основной пищи взрослых рыб этого рода.
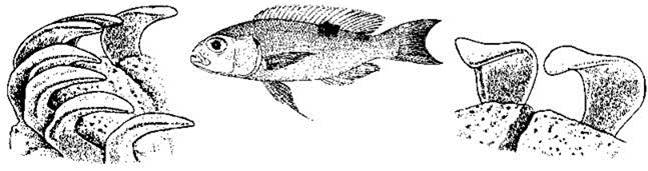
Рис. 1.31. Perissodus hecqui и его зубы в разных ракурсах. По Coulter (1991).
Таким образом, одновременно с образованием крупных озёр (или других больших по размеру водоёмов), ресурсы в которых резко отличаются от ресурсов в окружающих водоёмах, в них формируются экосистемы из имеющихся в данной местности. В результате некоторые таксоны преобразуются в эндемов. К аналогичным выводам пришёл и Я.И. Старобогатов (1970), анализируя пути формирования фауны моллюсков озера Танганьика. Время, необходимое для формирования новой экосистемы озера, в геологическом масштабе может быть ничтожно малым, небольшим даже по человеческим меркам. Из этого, в частности, следует, что эндемичная фауна Байкала — автохтонная, и образовалась она в самом Байкале, а большой возраст её видов, возможно, сильно преувеличен.