Биогеография общая и частная: суши, моря и континентальных водоёмов - 2017 год
Критерии вида - Виды, их критерии и причины видообразования - Био-экология, или синэкология
Понятие вида — одно из основных понятий биологии. Однако, хотя люди с совершенно различной подготовкой выделяют одни и те же виды, не существует простого операционного определения вида. Теоретико-философские концепции вида многочисленны, вероятно, их почти столько же, сколько желающих потеоретизировать на эту тему. Однако их значение для практической таксономии весьма невелико, в значительной степени эти теории являются “вещью в себе” и не предназначены для практического применения, поскольку они обычно не доведены до возможности их проверки на соответствие реальности. Ниже обсуждены операционные критерии вида, теория же затронута лишь в той степени, в которой она необходима для решения проблемы распознавания видов в природе.
Под видом обычно понимают “кирпичики” из которых состоит биосфера (популяционно-видовой уровень организации биосферы). Длительное время выделение видов основывалось почти исключительно на изучении морфологии. При этом вид понимали, как совокупность морфологически сходных особей. Такие совокупности полагали однородными внутри себя, сходно обособленными и стабильными, а задачей таксономии считали выявление таких дискретных совокупностей. Особей изучаемых выборок относили к разным видам, если обнаруживали существенные различия в их морфологии. Практическое применение этой концепции вида столкнулось с двумя проблемами: (1) изменчивостью и (2) тем, что отнюдь не всегда анализируемые совокупности особей можно было разбить на дискретные виды.
(1) Первоначальные интуитивные представления о виде как совокупности морфологически сходных особей с развитием генетики постепенно превратилось в требование панмиксии. Изменчивость стали признавать только географическую и индивидуальную, групповую изменчивость (кроме половой) считают в норме отсутствующей. Нарушение панмиксии считают единственно возможной причиной видообразования (аллопатрическое видообразование, видообразование в результате полиплоидии и т.п.). Симпатрическое существование или возникновение видов, не изолированных репродуктивно считают невозможным.
(2) Многие виды вполне дискретны и имеют ясные морфологические отличия ("хорошие виды”), но имеют в той или иной степени различающиеся морфотипы. Это ставит перед исследователем проблему: какую единицу из числа распознаваемых можно назвать видом. Также были обнаружены дискретные по тем или иным признакам совокупности, различия некоторых были незначительны или почти отсутствовали. Такие виды были названы Э. Майром sibling species (виды-двойники).
Обнаружение плохо различающихся видов привело к поиску дискретности в других признаках, помимо морфологических. Наиболее часто использовали элементы полового поведения: песни птиц, стрекотание прямокрылых, вспышки светлячков и т.п. Использовали также данные электрофореза белков, кариологии, цитологии, иммунологии и т. д. и т. п. Основная парадигма была лишь слегка модифицирована. Термин “вид” был дополнен терминами, которые были применимы к случаям плохо различимых видов: кроме термина “виды-двойники” появились термины “полувид”, “надвид” и т. п.
Развитие молекулярной биологии и генетики казалось бы позволяет решить проблему критерия вида. В основу генетической концепции положена та же идея о существовании обособленных стабильных видов, но критерием вида в этой концепции стали отличия геномов. Под видом стали понимать группу панмиктических особей, генетически изолированную от других таких групп. Разные особи одного вида при таком понимании вида имеют сходные ДНК, отличные от ДНК других видов. Сравнивая ДНК разных особей, можно оценить их сходство. В качестве критерия вида сторонники генетической концепции вида предлагают использовать генетическую дистанцию, измеряемую в процентах различия между парами гомологичных последовательностей ДНК. Проблема в том, что не только виды отличаются последовательностями ДНК, но и особи и популяции. Фактически изучение последовательностей ДНК даёт лишь информацию о степени сходства, но не ответ на вопрос о том, принадлежат сравниваемые особи к разным видам или к одному. Наиболее “экстремистски” настроенные сторонники генетической концепции вида считают возможным принять стандартное значение, превышение которого автоматически означает, что сравниваемые особи принадлежат к разным видам.
Генетическая концепция вида основана на теоретических представлениях, из которых наиболее очевидные следующие:
— генетическая дистанция между популяциями пропорциональна таксономическому рангу;
— избранные для сравнения гены изменяются с постоянной скоростью;
Практика применения в качестве критерия генетической дистанции показала, что:
— разные популяции одного вида могут различаться больше, чем хорошо различимые виды;
— один и тот же ген может эволюционировать с разной скоростью даже у близкородственных таксонов.
Примером несоответствия результатов генетической и морфологической концепций видов может служить исследование бурых Ursus arctos и белых U. maritimus медведей(Ceiridwen et al., 2011) показало, что, хотя белые медведи хорошо отличимы от бурых морфологией, биологией, а современные — и ядерными генными маркёрами, по мтДНК белые медведи образуют одну кладу с некоторыми популяциями бурых. Нет между этими видами и репродукционной изоляции: они способны скрещиваться, а их потомство плодовито (O'Harra, 2010). С последовательно генетических позиций правильно таксономически отразить эту ситуацию можно только разделив вид медведь бурый U. arctos на несколько, равноценных белому медведю. Такое решение очевидно неправильно.
Потомство многих видов в искусственных условиях (т. е. вне ЦС) вполне жизнеспособно, что показывает отсутствие у них действенной репродуктивной изоляции на уровне видов. Например, гибриды соболя (Martes zibellina) и куницы (M. martes) — кидусы — вполне плодовиты, что многократно проверено в пушном разведении. Ареалы соболя и куницы широко перекрываются (куница доходит на восток до Оби, а соболь на запад до Печоры), но самостоятельность их несомненна. Примеров видов, свободно скрещивающихся в неволе, но длительно сосуществующих в природе (т. е. в ЦС), очень много: все утки (Anatidae) и полорогие Bovidae, многие олени, их потомство вполне жизнеспособно, в случае уток это даже создаёт определённую проблему в зоопарках. Дополнительные примеры по позвоночным можно найти у С.С. Шварца (1980) и Е.Н. Панова (1989).
Весьма любопытная система, состоящая из трёх видов, один из которых — съедобная лягушка (Rana esculenta) — гибрид двух других: озёрной (R. ridibunda) и прудовой (R. lessonae) (Шабанов, Литвинчук, 2010). Виды этой системы свободно обмениваются генетической информацией и имеют различающиеся ареалы, в частности гибридный вид может обитать там, где нет одного или даже обоих родительских. Существование гибридного вида даёт возможность переноса через него генетической информации от одного родительского вида к другому и обратно. В результате, у большинства озёрных лягушек Западной Европы митохондриальная ДНК замещена митохондриальной ДНК прудовой. Т. е., если опираться лишь на митохондриальную ДНК, то пришлось бы признать, что R. ridibunda и R. lessonae — один вид.
Разнообразие механизмов таксонообразования у сосудистых растений значительно выше, чем у животных (Камелин, 2009). У растений широко распространены различные формы полиплоидии и создания видов в результате гибридизации ранее существовавших видов. Причём гибридизирующиеся виды могут принадлежать различным родам, и даже семействам. Обмен генетической информацией у них не ограничен половым процессом мужских и женских особей одного вида. Среди растений широко распространены и хорошо изучены генетиками межвидовые комплексы, объединяющие целые роды или большое количество видов в многовидовых родах. Виды таких комплексов потенциально геном-совместимы. В природе виды этих комплексов могут быть как вполне чёткими, так и плохо различимыми. Комплексы плохо различимых видов известны и среди животных, но они мало изучены генетиками. Кроме сомнительности представлений, положенных в основу генетического критерия вида, гораздо более важно, что вся эта теория применима только к видам с регулярным половым размножением. Однако виды, у которых половое размножение неизвестно с точки зрения таксономиста не отличаются от видов, размножающихся половым путём: у них сходны внутривидовые морфологическая изменчивость и уровень межвидовых различий (Fontaneto et al., 2007). Генетики решают эту проблему просто, рассматривая только виды, имеющие половое размножение, не замечая, что объяснение природы вида хотя и во многих, но отнюдь не во всех случаях делает теорию из общей — частной. Вопрос же о природе вида оставляют при этом открытым и подменяют другими. В генетическом критерии нет указания на то, почему в природе возникают виды. Генетика способна объяснить лишь механизмы видообразования, но не причины. Поэтому вопрос о причинах всячески стараются затушевать. Основной способ этого — объявить основной причиной видообразования изоляцию, а поддержание видами самостоятельности объяснять существованием изолирующих механизмов.
Многочисленные примеры отсутствия репродуктивной изоляции симпатричных длительно сосуществующих видов показывают, что широко распространённое мнение о её необходимости неверно. Несомненно, репродуктивная изоляция уменьшает потери популяции на неэффективное размножение. Многие исследователи из этого делают вывод, что виды не могут пойти на столь неэффективные потери. Однако было бы самонадеянным полагать, что мы лучше знаем, что выгодно, а что не выгодно видам. Если во многих случаях репродуктивная изоляция была показана, это вовсе не означает, что она должна быть ВСЕГДА. Более того, по крайней мере, в отдельных, но отнюдь не единичных случаях показано отсутствие репродуктивной изоляции. Если виды с этим мирятся, значит, с их точки зрения, потери невелики. Поэтому уже сейчас можно однозначно утверждать, что сама отсутствие репродуктивной изоляция не является критерием того, что данные дискретные совокупности особей не являются самостоятельными видами.
Не может служить критерием вида и наличие репродуктивной изоляции: виды могут образовывать симпатрические, но репродуктивно изолированные группы. Примером такого вида может служить ивовый листоед Lochamaea capreae (Северцов, 2008).
Показано, что у лягушек и птиц потомство межвидовых скрещиваний жизнеспособно даже у видов, разделившихся в самом начале кайнозоя, почти 60 млн. лет назад (Рис. 1.56). Из рисунка также видно, что возникновение репродуктивной изоляции является функцией не столько времени (накопления случайных различий генофондов, делающих их несовместимыми), а сколько способа размножения, т. е. способности видов переносить потери в результате межвидовых скрещиваний. Резюмируя можно сказать, что пытаться понять, что такое вид с помощью априорно выбранных критериев — бесперспективное занятие. Это можно сделать только поняв причины возникновения видов.
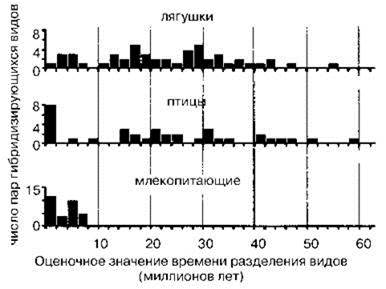
Рис. 1.56. Способность видов разных таксонов к гибридизации. При построении диаграммы учитывались только виды, способные к межвидовой гибридизации. По Zeh, Zeh (2000).