Биогеография общая и частная: суши, моря и континентальных водоёмов - 2017 год
Субклимаксы, экзогенная стабилизация ассоциаций - Сукцессионная система - Био-экология, или синэкология
Любая ассоциация может быть стабилизирована внешним воздействием. По механизмам возникновения С.М. Разумовский (1981) различал три типа субклимаксов: рецидивный, ретардационный и диаспорический.
Рецидивный субклимакс возникает при частом повторении одних и тех же нарушений, возвращающих ЦС в данном месте на уже пройденную сукцессионную стадию. При этом ни последовательность сериальных стадий, ни скорость протекания сукцессии не отличаются от нормальных, иными словами, рецидивный субклимакс лишен какой-либо качественной специфики. Это частный случай обычного явления “нарушение — восстановление”, отличающийся лишь значительной частотой нарушений. Частые нарушения создают лишь видимость стабильности, тогда как в действительности не происходит не только остановки, но даже замедления сукцессии.
Ретардационный субклимакс “возникает при крайнем замедлении смены под влиянием постоянно действующих факторов, не вызывающих нарушения ассоциации. Этот тип субклимакса известен лишь для экогенетических сукцессий и связан с внешним воздействием, замедляющими или останавливающим изменения биотопа, происходящее в результате экогенетической сукцессии. Например, тростниковые плавни в устьях наших рек неопределенно долго остаются на пионерной стадии евтрофной гидросерии, поскольку при достаточной скорости течения вода непрерывно уносит отлагаемый ими органический материал. При меньшей скорости течения торфообразование всё-таки происходит, но скорость смены в той или иной мере замедлена. В миниатюре эту же картину постепенного перехода от нормальной скорости смены до полной её остановки легко видеть на любой мелководной реке” (Разумовский, 1999: 281). Аналогичные процессы, по-видимому, имеют место в условиях повышенной гидродинамики в сообществах рыхлых грунтов. “Подобно рецидивному субклимаксу, ретардационный субклимакс не имеет качественной специфики, представляя собой доведённое до крайности весьма обычное явление замедления экогенетической сукцессии. Последовательность смены при этом остается нормальной, а уменьшение её скорости может иметь самую разнообразную величину” (Разумовский, 1999: 281).
Сходную картину могут дать и постоянные селективные внешние воздействия. Например, в березняках, выросших в результате демутационной сукцессии (Рис. 1.37) возобновлению ели могут препятствовать рекреационная нагрузка или выпас скота, уничтожающего подрост ели.
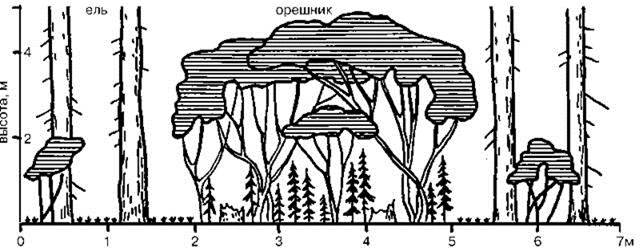
Рис. 1.42. Орешниково-еловый диаспорический субклимакс на севере Смоленской области. По Разумовскому (1981).
Диаспорический субклимакс. В отличие от рецидивного и ретардационного субклимаксов здесь нет внешнего воздействия, препятствующего прохождению сукцессии. Этот тип субклимакса возникает, если виды следующей сериальной ассоциации по какой-либо причине не могут попасть в данное место, а, следовательно, не может сформироваться следующая сериальная ассоциация. С.М. Разумовский приводит пример диаспорического субклимакса на севере Смоленской области в верховьях Днепра. Эта “обширная территория, совершенно лишенная дуба и волосистой осоки (Carexpilosa). Отсутствие дуба вызвано там не деятельностью человека, а чрезмерной экологической однородностью территории. Плоская равнина, тянущаяся на десятки километров, сплошь занята сообществами евтрофной гидросерии, неприемлемыми для дуба. Небольшие участки этой равнины, на которых экогенез уже достиг стадии климакса, отделены от ближайших дубрав слишком большими заболоченными пространствами, преодолеть которые миграция дуба еще не смогла, хотя расселение орешника на новообразовавшиеся участки климакса уже происходит... В ельнике- кисличнике возникает полог орешника... После распада елового древостоя орешник быстро приобретает древовидную форму (за счет роста одного из побегов плейохазия и подавления остальных) и достигает высоты 10—12 м при диаметре на высоте груди 18—20 см. В образующемся орешниковом лесу с покровом кислицы обильно возобновляется ель. Как только новый еловый древостой перерастет по высоте древостой орешника, последний быстро выпадает, и сообщество снова превращается в ельник- кисличник, чтобы снова начать тот же цикл” (Разумовский, 1981: 38—39) (Рис. 1.42). Данный тип субклимаксов, по-видимому, обычный в наземных ЦС, в морских, если и наблюдают, то крайне редко из-за гораздо большей способности морских организмов к расселению.
К трём вышеперечисленным типам следует добавить ещё один, который наблюдают в тех случаях, когда условия на данной площади выходят за пределы экологической амплитуды данной ЦС, но нет других ЦС, что смогли бы заселить данный участок. Такого типа субклимаксные ассоциации характерны для распреснённых районов эстуариев. Поскольку климакс в данном случае не достигается из-за отличий биотопа, которые не могут быть устранены действием ЦС, я предлагаю назвать такой субклимакс биотопическим.
Биотопическим субклимаксом можно считать ранние ксеросериальные стадии, широко распространившиеся на Земле в результате эрозии почв (пустыни, полупустыни, саванны и т. п., подробнее см. Главу 3). В девственных ЦС биогены хранились в деревьях (лесные ассоциации) или почве (безлесные ассоциации). Сведение лесов и эрозия почв приводит к утрате биогенов. Это препятствует прохождению сукцессий далее ранних ксеросериальных стадий. Повторное накопление биогенов, если и возможно, потребует многих тысячелетий.
Если причины, по которым возникают субклимаксы, обычны в течение столь длительного времени, что их можно рассматривать как фактор внешней среды, субклимаксные ассоциации постепенно эволюируют в климаксные, т. е. происходит формирование новой ЦС. Если же условия меняются непрерывно, то столь же непрерывно происходит и изменение сообществ, как это можно наблюдать в Аральском и Чёрном морях.