Биогеография общая и частная: суши, моря и континентальных водоёмов - 2017 год
Сукцессионная система - что это такое - Сукцессионная система - Био-экология, или синэкология
Итак, многочисленные примеры показывают, что в некоторых случаях быстро (иногда практически мгновенно) образуются эндемичные виды и экосистемы. Поскольку образование новых видов может происходить весьма быстро, кажется вероятным, что формирование специфических взаимодействий между видами может происходить по крайней мере столь же стремительно, а, вероятно, и ещё быстрее. Закономерности этого процесса, т. е. эволюция, возникновение и развитие экосистем, и составляют основное содержание концепции структурализма. Парадигма структурализма сравнительно малоизвестна. Её изложения нет ни в одном учебнике или монографии как в отечественных (если не считать фрагменты положений, встречающиеся в трудах континуалистов), так и, насколько мне известно, в иностранных. В то же время, по моему мнению, она адекватнее описывает структуру биосферы. Поэтому ниже, параллельно с обсуждением общих вопросов экологии, дано изложение этой парадигмы. Желающие более углублённо ознакомиться с теорией структурализма могут обратиться к сборникам трудов, изданных издательством КМК (Разумовский, 1999, 2011; Жерихин, 2003).
Размеры экосистем, возникающих в представленных выше примерах, гораздо больше размеров отдельных биоценозов и сообществ. Фактически многие из таких экосистем образованы целым комплексом сообществ. Поэтому для таких крупномасштабных экосистем F. Clements предложил термин биом. Однако за последнее время смысл термина “биом”, особенно в русскоязычной литературе, становился всё более расплывчатым. При этом “биомы” выделяют по признакам, которые автор считает существенными и у разных авторов эти признаки не совпадают. Не удивительно, что в результате у разных авторов не только смысл отдельных биомов, но и их число могут сильно различаться. В то же время о сукцессионных изменениях даже не упоминают. Поэтому С.М. Разумовским был предложен новый термин для того же явления — ценотическая, или сукцессионная система (Разумовский, 1969; Киселева, Разумовский Расницын, 1969; Киселева, Разумовский, 1971; и др.), в дальнейшем — СС, или ЦС*.
Новая ЦС формируeтся, когда возникает пространство:
— На котором не может эффективно функционировать ни одна из ЦС, могущих туда попасть.
— Достаточно большое для существования ЦС и с предсказуемыми условиями.
— Существующее на протяжении времени, достаточного для формирования ЦС.
Данные условия могут показаться неконкретными и слишком строгими, а соответствующие им площади — редко встречающимися в природе. В действительности это не так: подобные площади возникают постоянно, и всегда мы наблюдаем возникновение ЦС и обычно сопровождающий их очаг эндемизма (видов, ареалы которых ограничены только данным районом). Очевидно, что примерами подобных площадей являются:
— некоторые крупные и не очень крупные озёра (Байкал, Ньяса и т. п.), а также острова;
— достаточно крупные горы или горные системы посреди равнин;
— ультраабиссальные желоба посреди абиссальных равнин;
— изолированные глубоководные котловины (Северный Ледовитый океан, Японское и Средиземное моря);
Сложнее заметить, что этим условиям полностью удовлетворяют:
— высокие широты, после произошедшего в олигоцене — миоцене увеличения температурного градиента между экватором и полюсом (относится в равной степени к территориям и акваториям): в конце палеоцена поверхностная температура воды в Антарктике достигала 18° С.
— приэкваториальные районы, после произошедшего в то же время повышения температуры выше экологической амплитуды существовавших в этих местах ранее субтропических СС.
— вертикальные зоны на суше и, особенно, в океане.
Ни в одном из приведённых выше примеров (а число их можно значительно увеличить) не возникала преграда. Ареал формирования новой ЦС всегда оставался связанным с ареалом материнских ЦС. В последних трёх случаях формирующиеся ЦС имели общие границы с ЦС-донорами на протяжении тысяч километров! Поэтому Рис. 1.16 не то чтобы не верен, но подчёркивает второстепенную, совершенно не обязательную причину видообразования — фрагментацию ранее единой биоты. При этом действительно важная причина, имеющая гораздо большее значение и распространение, — возникновение на обширных площадях условий, выходящих за пределы, в которых существующие ЦС способны нормально функционировать, — на нём никак не отражена. По-видимому, более правильной следует считать совершенно другие схемы возникновения биот и видов, изображённые на Рис. 1.34 и Рис. 1.35. Наличие трудно преодолимых преград, обуславливающих границы между ЦС, которые обычно считают непременным условием существования биогеографических границ, в действительности — редчайшее исключение, по крайней мере, в наземных и донных ЦС. Такие преграды обычны в пелагиали, но ситуация здесь принципиально иная (подробнее см. Главу 6). Пресноводные же ЦС обычно фрагментированы и фактически аналогичны изолированным островам. Но и здесь непреодолимые преграды полностью отсутствуют при формировании ЦС крупных озёр (подробнее см. Главу 4). Поэтому широко распространённое мнение, что эндемизм невозможен без изоляции неверно. Оно основано на некритической экстраполяции данных по островным ЦС на гораздо более распространённую ситуацию непрерывного распространения. Начало этим экстраполяциям было положено Эрнстом Майром, Ph.D. диссертация которого посвящена птицам островов Полинезии.
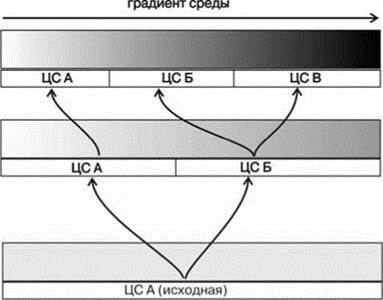
Рис. 1.34. Возникновение СС и биот с позиции структурализма.
Обратите внимание на то, что СС формируются на отрезках плавного градиента. В отличие от рис. 1.15, на схеме полностью отсутствуют какие-либо преграды между формирующимися сс. Нет даже областей увеличенных градиентов. Границы между СС тем не менее резкие. Дополнительные пояснения в тексте.
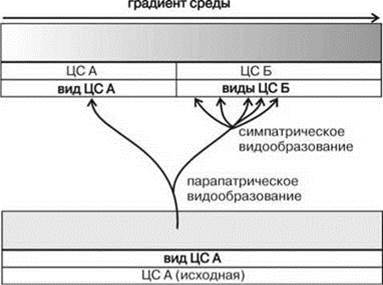
Рис. 1.35. Видообразование с позиции структурализма.
Обратите внимание на то, что виды возникают сначала парапатрически на отрезках плавного градиента при возникновении двух биот. При формировании новой биоты происходит вспышка симпатрического видообразования. Дополнительные пояснения в тексте.
При образовании ЦС вид должен встроиться в формирующиеся сообщества, а не приспособиться к некоей абстрактной неизменной внешней среде. Биологические свойства большинства видов (потенциальная ниша) позволяют им использовать в различных ассоциациях разные ресурсы. Если старые виды не способны эффективно использовать имеющиеся на территории данной ЦС ресурсы, образуются новые виды.
Итак, ЦС возникают на достаточно однородных площадях, где условия выходят за пределы экологических амплитуд ЦС, которые могут на эту площадь попасть. Существование какого-либо барьера при этом вовсе не обязательно, вероятно, его обычно и нет — ареал формирующейся ЦС остаётся широко сообщающимся с ареалами старых ЦС. Но, поскольку условия в ареале формирующейся ЦС существенно отличаются от условий в ареалах старых, между их ареалами существует градиент условий. Граница между ЦС устанавливается в области таких значений внешних факторов, при которых конкурентоспособность (способность занять и удерживать территорию) контактирующих ЦС равна.
Понятие ЦС позволяет достаточно чётко разграничить экологию и биогеографию. Границу между ними логично провести по уровню ЦС: объекты ранга ЦС и ниже — предмет экологии; распределение ЦС, их взаимодействие и эволюция, а также сравнение ЦС и их частей по разным показателям (т. е. изучение явлений в масштабе большем, чем ЦС) — предмет биогеографии.
Свойства конкретной ЦС не сводимы к сумме свойств видов, входящих в неё. ЦС — это другой уровень организации, и в этом смысле её можно назвать квазиорганизмом.
Биосфера поделена на сравнительно небольшое число ЦС. На суше — 174 ЦС (Разумовский, 1999), на морском дне по моим подсчётам — около 300, в открытых водах океана — около 60, что в сумме даёт около 600. Число ЦС в пресных водах из- за их сильной фрагментированности и сравнительно слабой оформленности оценить наиболее сложно. В качестве оценки можно принять число выделенных экорегионов (Abell et al., 2008) — 830. Число пресноводных ЦС, возможно, немного больше, поскольку в крупных озёрах, которые были приняты за один экорегион, ЦС несколько. Впрочем, возможно, что некоторые ЦС разделены на несколько экорегионов. Вероятное число пресноводных ЦС — 500—1000 — как минимум половина ЦС Земли, но структурно они развиты в наименьшей степени.
ЦС используют энергию, поступающую в их ареал. Низший трофический уровень существенного числа ЦС представлен гетеротрофами, потребляющими поступающее в ареал ЦС органическое вещество в форме детрита. Такие лишённые автотрофов ЦС занимают большую часть поверхности Земли — морское дно за пределами фитали и толщу воды глубже главного термоклина (1—1,5 км).
В.Д. Фёдоров и Т.Г. Гильманов (1980: 38) предлагают два критерия экосистем: отсутствие внутренних причин, ограничивающих (1) размеры и (2) время существования. Объекты, удовлетворяющие этим условиям, они считают экосистемами, не удовлетворяющие — их частями. Первое требование следует считать субъективным, а потому избыточным, вполне достаточно только второго: экосистемами следует считать только сообщества, способные удерживать занятую ими территорию неограниченно долго. Очевидно, что ему удовлетворяют только ЦС, и не удовлетворяют различные сообщества, обычно (в том числе и указанными авторами) рассматриваемые как экосистемы: березняки, сосняки, речушки и т. д. и т. п., поскольку время их существование ограничено скоростью прохождения сукцессий. Даже климаксные ассоциации, взятые отдельно, нельзя считать экосистемами, хотя теоретически время их существования неограниченно. В реальности же рано или поздно экзогенное воздействие разрушит их, но их восстановление невозможно без прохождения сукцессий, т. е. возникновения в этом месте других сообществ. Необходимо рассматривать экосистемы не статично, а учитывая их динамику, не как фотографии, а как кино. Это как семейная жизнь: свадьба и шлепанцы — это лишь разные её временные состояния.